

Dependence of tumor cell lines and patient-derived tumors on the NAD salvage pathway renders them sensitive to NAMPT inhibition with GNE-618
- PMID: 24204194
- PMCID: PMC3819631
- DOI: 10.1593/neo.131304
Dependence of tumor cell lines and patient-derived tumors on the NAD salvage pathway renders them sensitive to NAMPT inhibition with GNE-618
Abstract
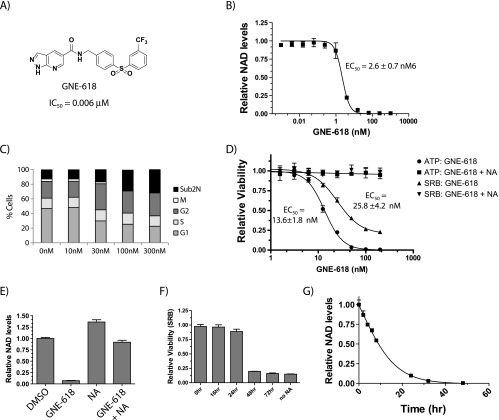
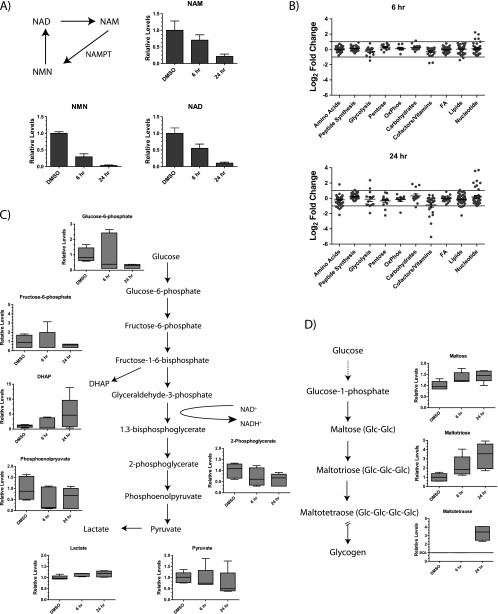
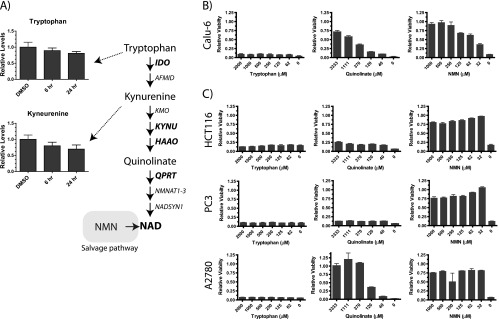
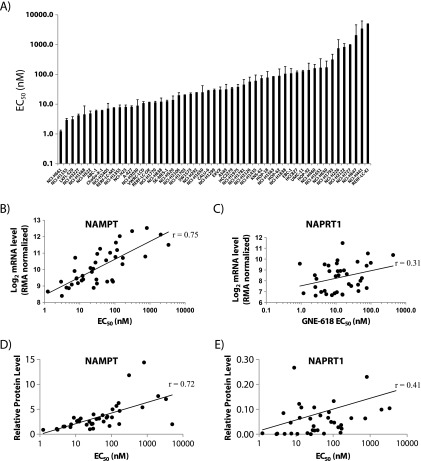
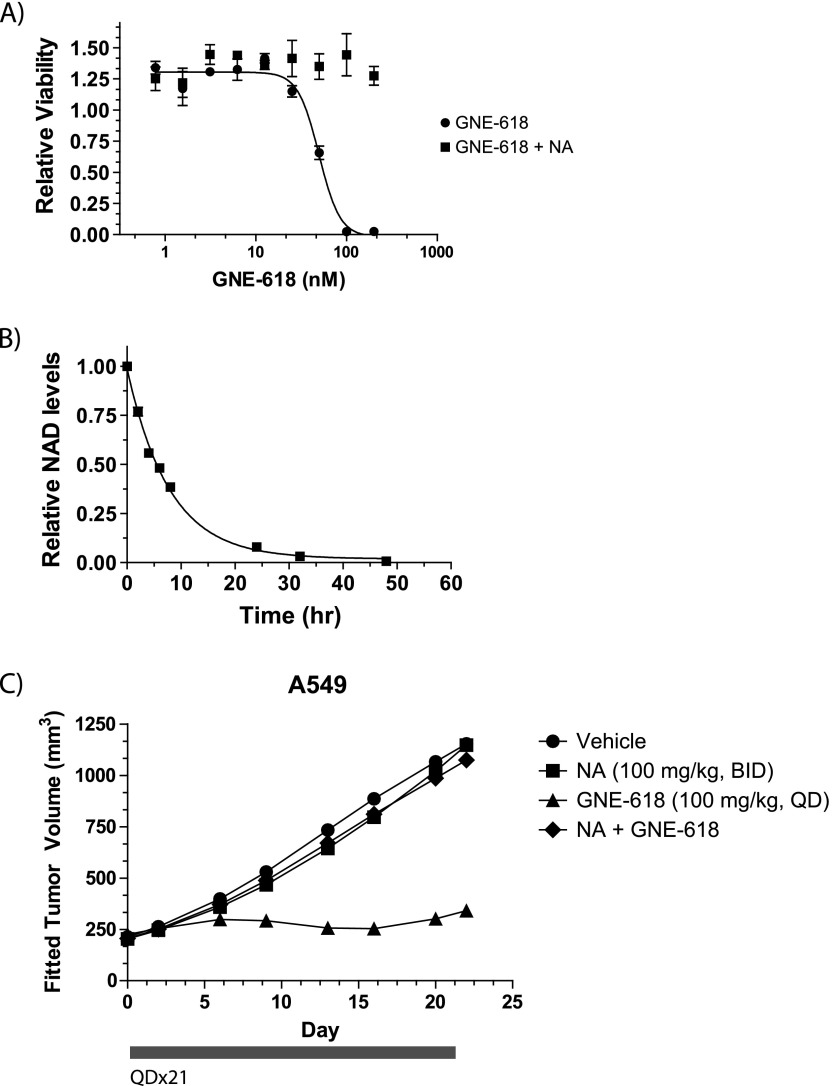
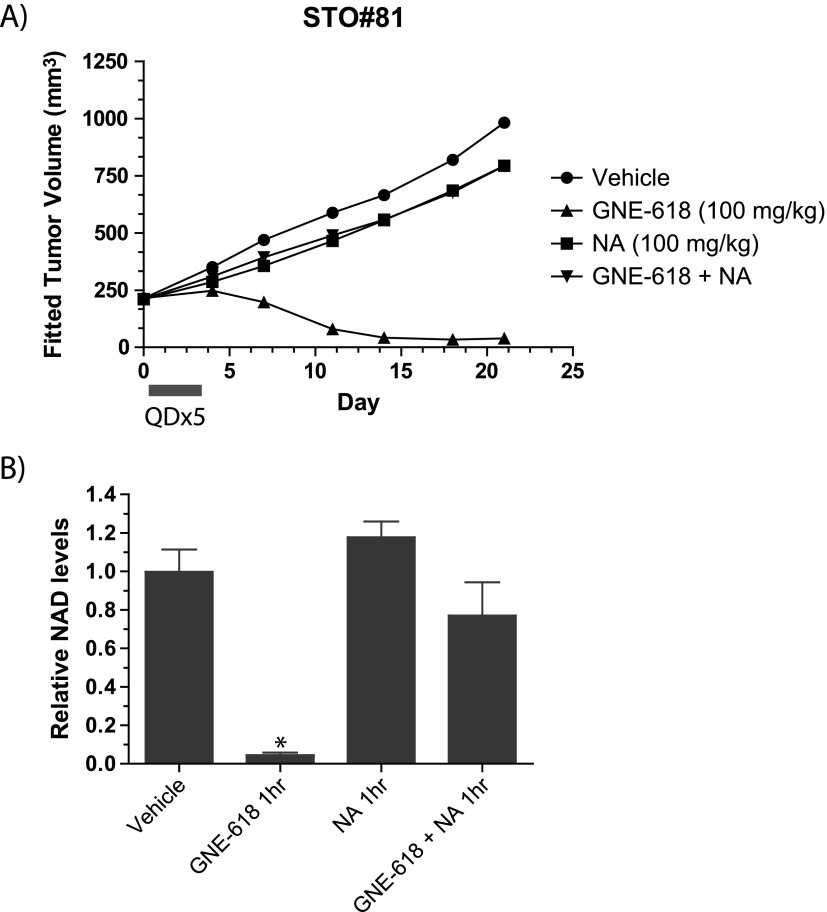
Nicotinamide adenine dinucleotide (NAD) is a critical metabolite that is required for a range of cellular reactions. A key enzyme in the NAD salvage pathway is nicotinamide phosphoribosyl transferase (NAMPT), and here, we describe GNE-618, an NAMPT inhibitor that depletes NAD and induces cell death in vitro and in vivo. While cells proficient for nicotinic acid phosphoribosyl transferase (NAPRT1) can be protected from NAMPT inhibition as they convert nicotinic acid (NA) to NAD independent of the salvage pathway, this protection only occurs if NA is added before NAD depletion. We also demonstrate that tumor cells are unable to generate NAD by de novo synthesis as they lack expression of key enzymes in this pathway, thus providing a mechanistic rationale for the reliance of tumor cells on the NAD salvage pathway. Identifying tumors that are sensitive to NAMPT inhibition is one potential way to enhance the therapeutic effectiveness of an NAMPT inhibitor, and here, we show that NAMPT, but not NAPRT1, mRNA and protein levels inversely correlate with sensitivity to GNE-618 across a panel of 53 non-small cell lung carcinoma cell lines. Finally, we demonstrate that GNE-618 reduced tumor growth in a patient-derived model, which is thought to more closely represent heterogeneous primary patient tumors. Thus, we show that dependence of tumor cells on the NAD salvage pathway renders them sensitive to GNE-618 in vitro and in vivo, and our data support further evaluation of the use of NAMPT mRNA and protein levels as predictors of overall sensitivity.
Figures
References
-
- Tomlinson IP, Alam NA, Rowan AJ, Barclay E, Jaeger EE, Kelsell D, Leigh I, Gorman P, Lamlum H, Rahman S, et al. Germline mutations in FH predispose to dominantly inherited uterine fibroids, skin leiomyomata and papillary renal cell cancer. Nat Genet. 2002;30(4):406–410. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous