

PNUTS/PP1 regulates RNAPII-mediated gene expression and is necessary for developmental growth
- PMID: 24204300
- PMCID: PMC3814315
- DOI: 10.1371/journal.pgen.1003885
PNUTS/PP1 regulates RNAPII-mediated gene expression and is necessary for developmental growth
Abstract
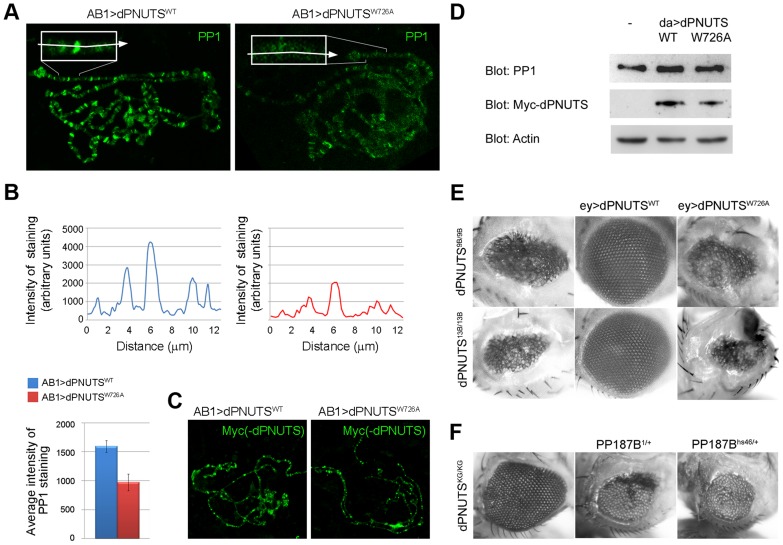
In multicellular organisms, tight regulation of gene expression ensures appropriate tissue and organismal growth throughout development. Reversible phosphorylation of the RNA Polymerase II (RNAPII) C-terminal domain (CTD) is critical for the regulation of gene expression states, but how phosphorylation is actively modified in a developmental context remains poorly understood. Protein phosphatase 1 (PP1) is one of several enzymes that has been reported to dephosphorylate the RNAPII CTD. However, PP1's contribution to transcriptional regulation during animal development and the mechanisms by which its activity is targeted to RNAPII have not been fully elucidated. Here we show that the Drosophila orthologue of the PP1 Nuclear Targeting Subunit (dPNUTS) is essential for organismal development and is cell autonomously required for growth of developing tissues. The function of dPNUTS in tissue development depends on its binding to PP1, which we show is targeted by dPNUTS to RNAPII at many active sites of transcription on chromosomes. Loss of dPNUTS function or specific disruption of its ability to bind PP1 results in hyperphosphorylation of the RNAPII CTD in whole animal extracts and on chromosomes. Consistent with dPNUTS being a global transcriptional regulator, we find that loss of dPNUTS function affects the expression of the majority of genes in developing 1(st) instar larvae, including those that promote proliferative growth. Together, these findings shed light on the in vivo role of the PNUTS-PP1 holoenzyme and its contribution to the control of gene expression during early Drosophila development.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
-
- Phatnani HP, Greenleaf AL (2006) Phosphorylation and functions of the RNA polymerase II CTD. Genes Dev 20: 2922–2936. - PubMed
-
- Bataille AR, Jeronimo C, Jacques PE, Laramee L, Fortin ME, et al. (2012) A universal RNA polymerase II CTD cycle is orchestrated by complex interplays between kinase, phosphatase, and isomerase enzymes along genes. Mol Cell 45: 158–170. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
