

Glutaredoxins are essential for stress adaptation in the cyanobacterium Synechocystis sp. PCC 6803
- PMID: 24204369
- PMCID: PMC3816324
- DOI: 10.3389/fpls.2013.00428
Glutaredoxins are essential for stress adaptation in the cyanobacterium Synechocystis sp. PCC 6803
Abstract
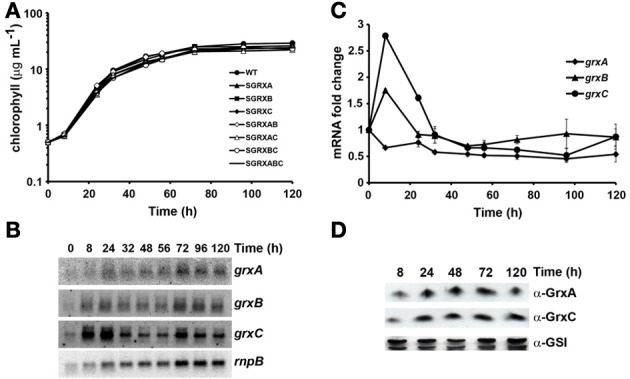
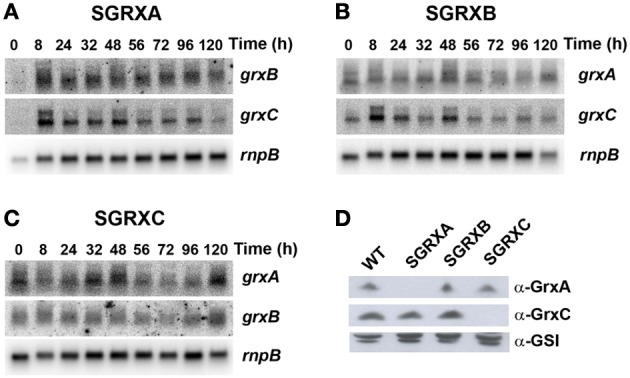
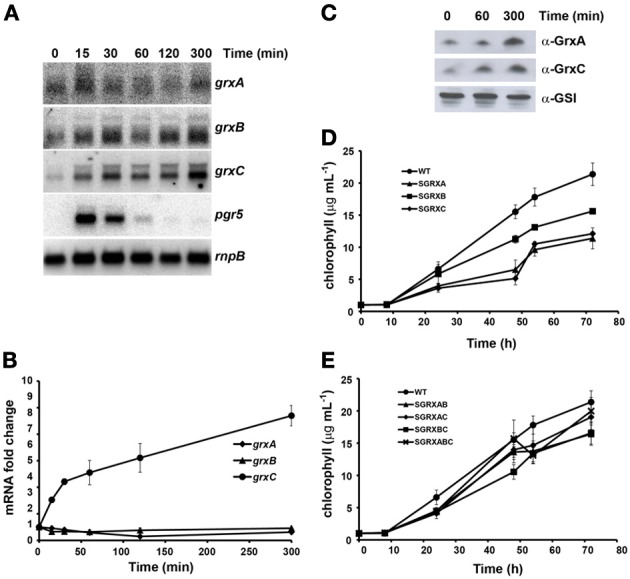
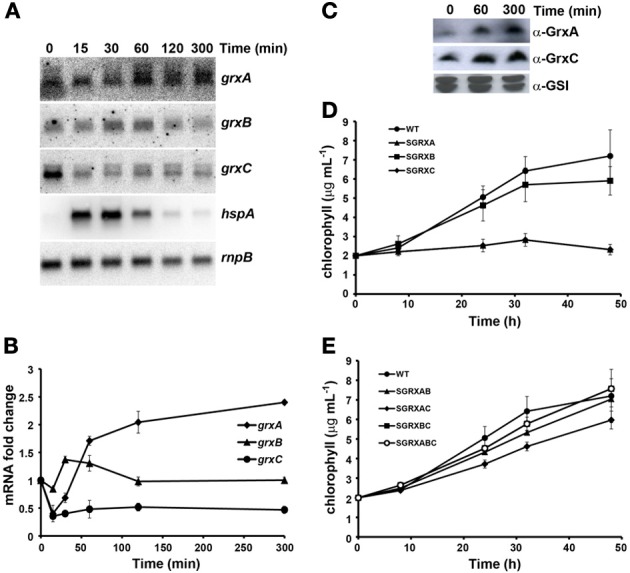
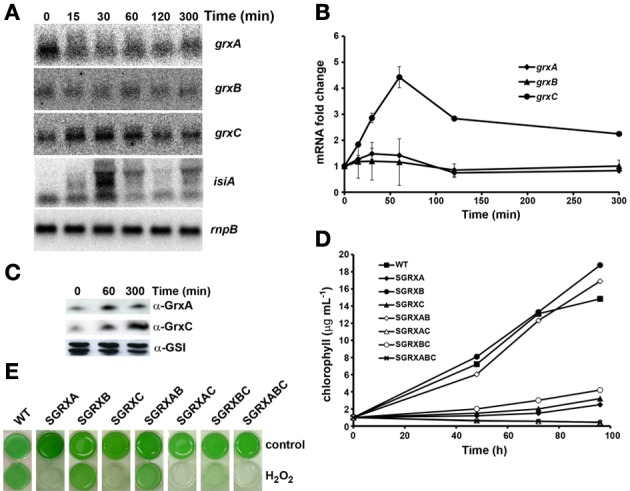
Glutaredoxins are small redox proteins able to reduce disulfides and mixed disulfides between GSH and proteins. Synechocystis sp. PCC 6803 contains three genes coding for glutaredoxins: ssr2061 (grxA) and slr1562 (grxB) code for dithiolic glutaredoxins while slr1846 (grxC) codes for a monothiolic glutaredoxin. We have analyzed the expression of these glutaredoxins in response to different stresses, such as high light, H2O2 and heat shock. Analysis of the mRNA levels showed that grxA is only induced by heat while grxC is repressed by heat shock and is induced by high light and H2O2. In contrast, grxB expression was maintained almost constant under all conditions. Analysis of GrxA and GrxC protein levels by western blot showed that GrxA increases in response to high light, heat or H2O2 while GrxC is only induced by high light and H2O2, in accordance with its mRNA levels. In addition, we have also generated mutants that have interrupted one, two, or three glutaredoxin genes. These mutants were viable and did not show any different phenotype from the WT under standard growth conditions. Nevertheless, analysis of these mutants under several stress conditions revealed that single grxA mutants grow slower after H2O2, heat and high light treatments, while mutants in grxB are indistinguishable from WT. grxC mutants were hypersensitive to treatments with H2O2, heat, high light and metals. A double grxAgrxC mutant was found to be even more sensitive to H2O2 than each corresponding single mutants. Surprisingly a mutation in grxB suppressed totally or partially the phenotypes of grxA and grxC mutants except the H2O2 sensitivity of the grxC mutant. This suggests that grxA and grxC participate in independent pathways while grxA and grxB participate in a common pathway for H2O2 resistance. The data presented here show that glutaredoxins are essential for stress adaptation in cyanobacteria, although their targets and mechanism of action remain unidentified.
Keywords: cyanobacteria; glutaredoxin; heat shock; high light; metal resistance; oxidative stress; redox regulation; stress.
Figures

Similar articles
-
Antioxidant ability of glutaredoxins and their role in symbiotic nitrogen fixation in Rhizobium leguminosarum bv. viciae 3841.Appl Environ Microbiol. 2021 Mar 1;87(4):e01956-20. doi: 10.1128/AEM.01956-20. Epub 2020 Dec 4. Appl Environ Microbiol. 2021. PMID: 33277272 Free PMC article.
-
The glutathione/glutaredoxin system is essential for arsenate reduction in Synechocystis sp. strain PCC 6803.J Bacteriol. 2009 Jun;191(11):3534-43. doi: 10.1128/JB.01798-08. Epub 2009 Mar 20. J Bacteriol. 2009. PMID: 19304854 Free PMC article.
-
Characterization of Escherichia coli null mutants for glutaredoxin 2.J Biol Chem. 2002 Mar 29;277(13):10861-8. doi: 10.1074/jbc.M111024200. Epub 2001 Dec 10. J Biol Chem. 2002. PMID: 11741965
-
Universal Molecular Triggers of Stress Responses in Cyanobacterium Synechocystis.Life (Basel). 2019 Aug 20;9(3):67. doi: 10.3390/life9030067. Life (Basel). 2019. PMID: 31434306 Free PMC article. Review.
-
Glutaredoxins: glutathione-dependent redox enzymes with functions far beyond a simple thioredoxin backup system.Antioxid Redox Signal. 2004 Feb;6(1):63-74. doi: 10.1089/152308604771978354. Antioxid Redox Signal. 2004. PMID: 14713336 Review.
Cited by
-
Responses to oxidative and heavy metal stresses in cyanobacteria: recent advances.Int J Mol Sci. 2014 Dec 31;16(1):871-86. doi: 10.3390/ijms16010871. Int J Mol Sci. 2014. PMID: 25561236 Free PMC article. Review.
-
Ancestors in the Extreme: A Genomics View of Microbial Diversity in Hypersaline Aquatic Environments.Results Probl Cell Differ. 2024;71:185-212. doi: 10.1007/978-3-031-37936-9_10. Results Probl Cell Differ. 2024. PMID: 37996679
-
Genomic Analysis of Serratia plymuthica MBSA-MJ1: A Plant Growth Promoting Rhizobacteria That Improves Water Stress Tolerance in Greenhouse Ornamentals.Front Microbiol. 2021 May 11;12:653556. doi: 10.3389/fmicb.2021.653556. eCollection 2021. Front Microbiol. 2021. PMID: 34046022 Free PMC article.
-
Antioxidant ability of glutaredoxins and their role in symbiotic nitrogen fixation in Rhizobium leguminosarum bv. viciae 3841.Appl Environ Microbiol. 2021 Mar 1;87(4):e01956-20. doi: 10.1128/AEM.01956-20. Epub 2020 Dec 4. Appl Environ Microbiol. 2021. PMID: 33277272 Free PMC article.
-
Thiol-based redox homeostasis and signaling.Front Plant Sci. 2014 Jun 10;5:266. doi: 10.3389/fpls.2014.00266. eCollection 2014. Front Plant Sci. 2014. PMID: 24959171 Free PMC article. No abstract available.
References
-
- Allakhverdiev S. I., Nishiyama Y., Miyairi S., Yamamoto H., Inagaki N., Kanesaki Y., et al. (2002). Salt stress inhibits the repair of photodamaged photosystem II by suppressing the transcription and translation of psbA genes in synechocystis. Plant Physiol. 130, 1443–1453 10.1104/pp.011114 - DOI - PMC - PubMed
-
- Bedhomme M., Adamo M., Marchand C. H., Couturier J., Rouhier N., Lemaire S. D., et al. (2012). Glutathionylation of cytosolic glyceraldehyde-3-phosphate dehydrogenase from the model plant Arabidopsis thaliana is reversed by both glutaredoxins and thioredoxins in vitro. Biochem. J. 445, 337–347 10.1042/BJ20120505 - DOI - PubMed
-
- Belli G., Molina M. M., Garcia-Martinez J., Perez-Ortin J. E., Herrero E. (2004). Saccharomyces cerevisiae glutaredoxin 5-deficient cells subjected to continuous oxidizing conditions are affected in the expression of specific sets of genes. J. Biol. Chem. 279, 12386–12395 10.1074/jbc.M311879200 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases