

Calcium/calcineurin synergizes with prostratin to promote NF-κB dependent activation of latent HIV
- PMID: 24204950
- PMCID: PMC3813743
- DOI: 10.1371/journal.pone.0077749
Calcium/calcineurin synergizes with prostratin to promote NF-κB dependent activation of latent HIV
Abstract
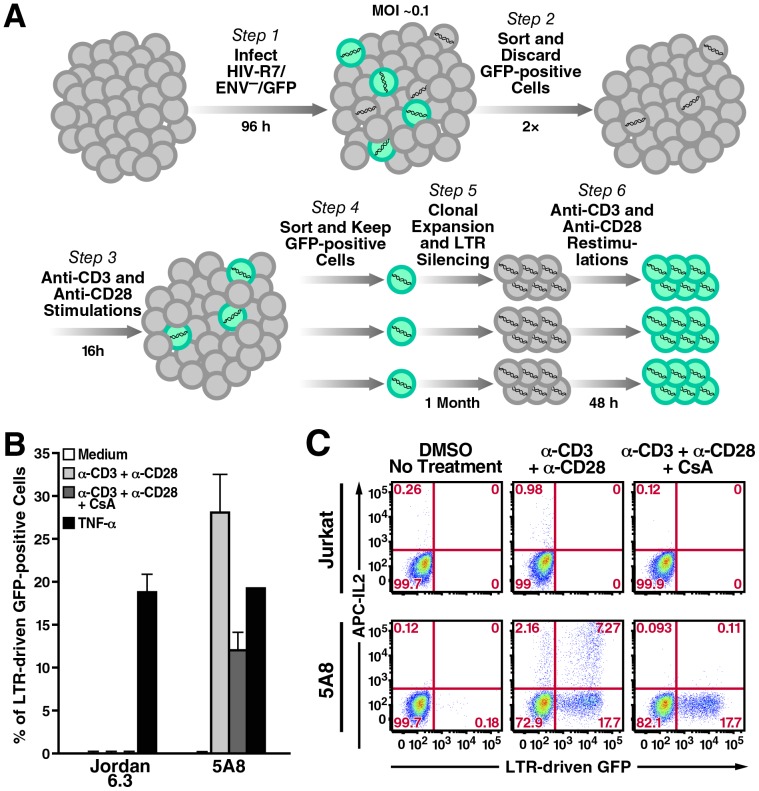
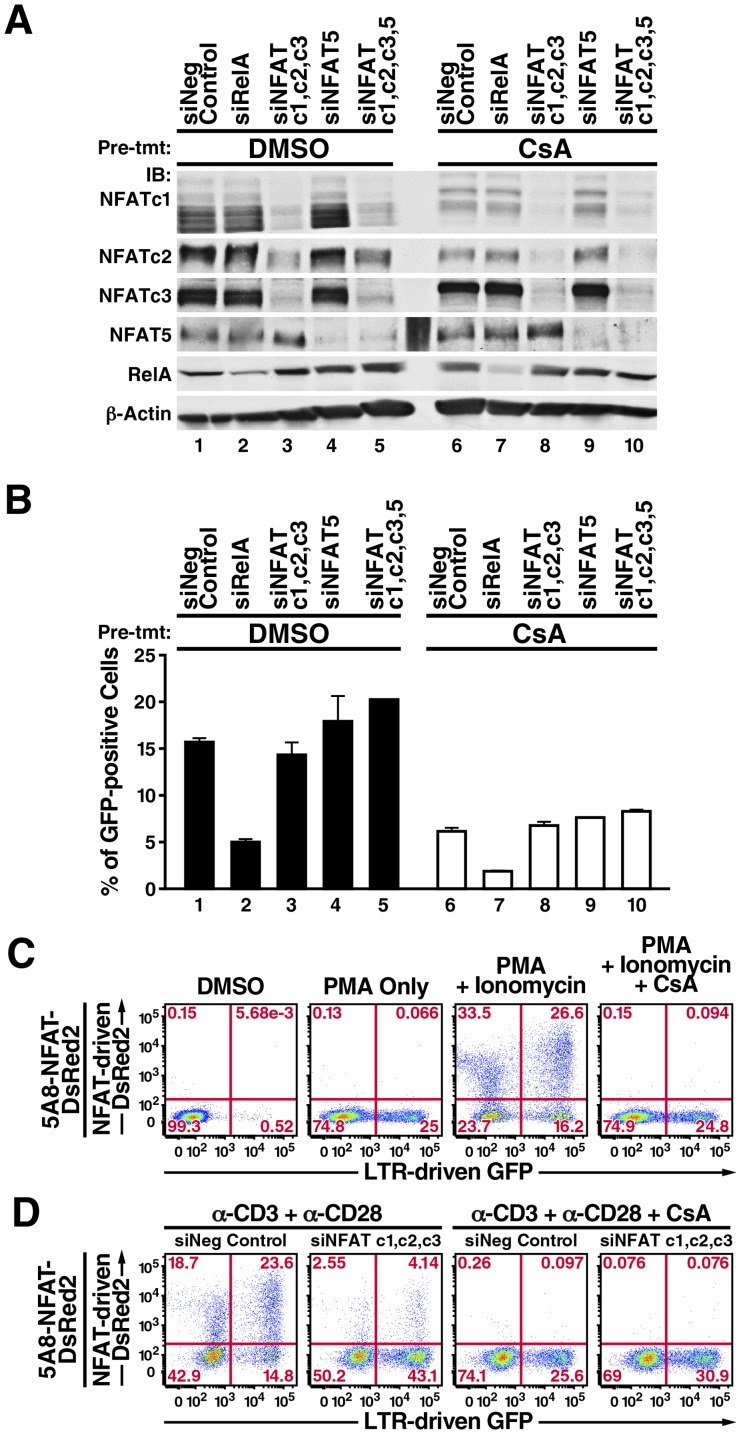
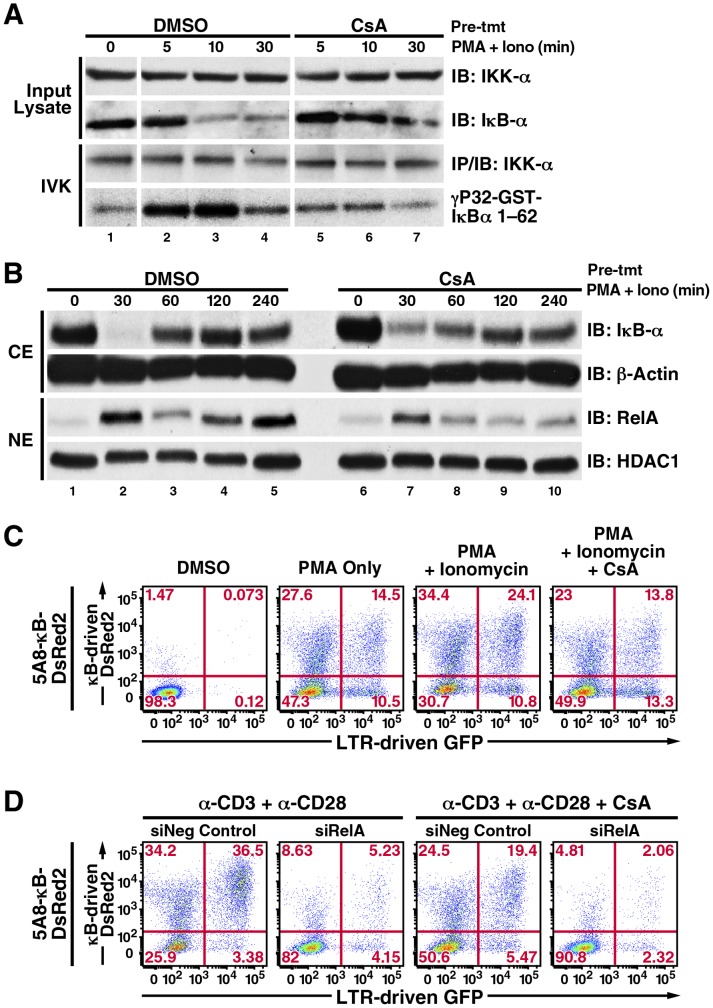
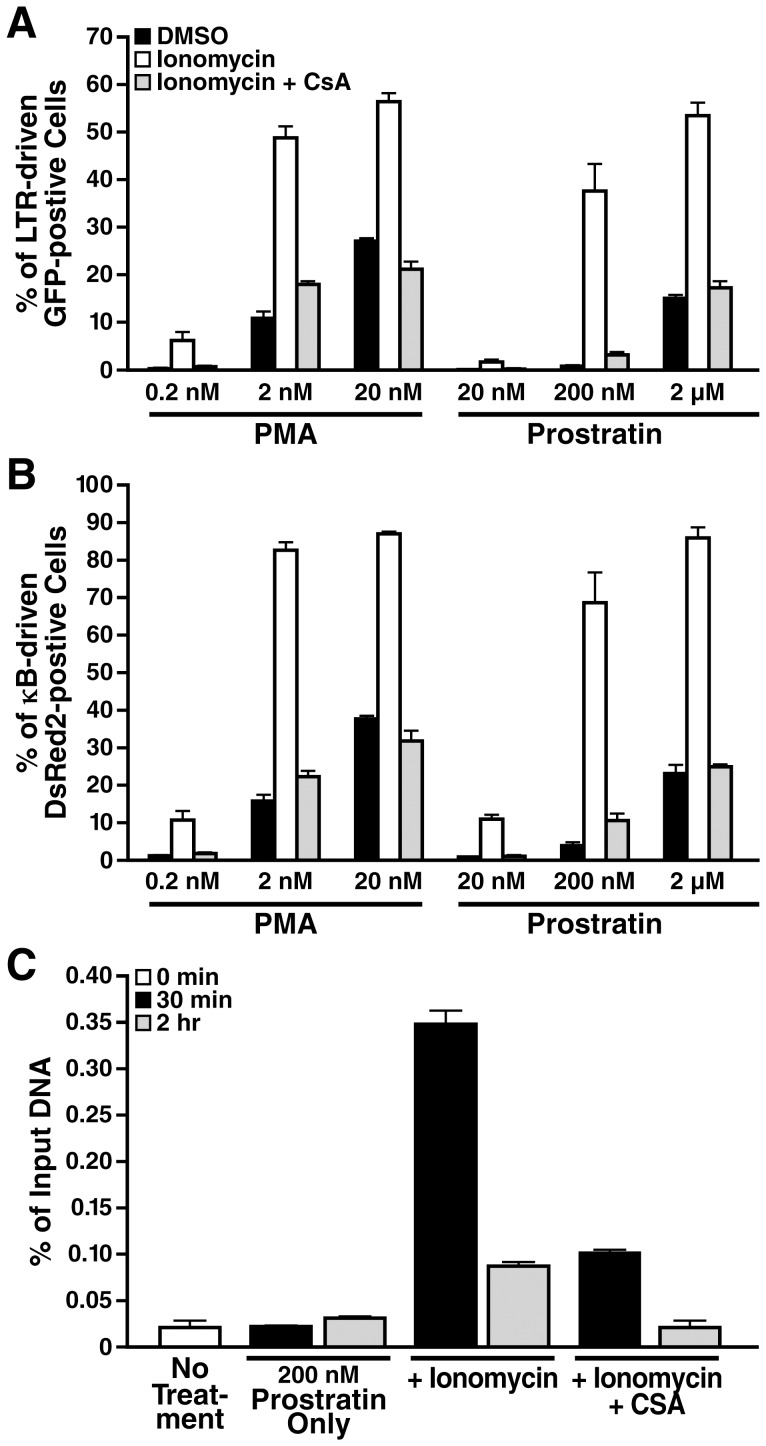
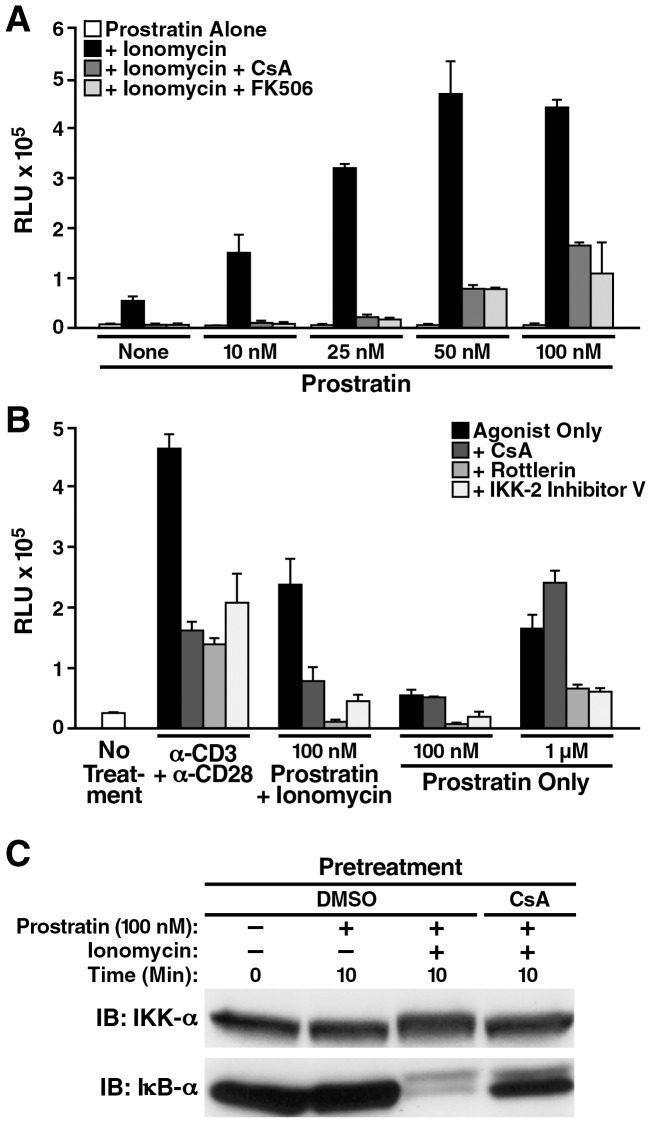
Attempts to eradicate HIV have been thwarted by the persistence of a small pool of quiescent memory CD4 T cells that harbor a transcriptionally silent, integrated form of the virus that can produce infectious virions following an anamnestic immune response. Transcription factors downstream of T-cell receptor activation, such as NF-κB/Rel and nuclear factor of activated T cells (NFAT) transcription members, are considered important regulators of HIV transcription during acute HIV infection. We now report studies exploring their precise role as antagonists of HIV latency using cell and primary CD4 T cell models of HIV-1 latency. Surprisingly, RNA interference studies performed in J-Lat CD4 T cells suggested that none of the NFATs, including NFATc1, NFATc2, NFATc3, and NFAT5, played a key role in the reactivation of latent HIV. However, cyclosporin A markedly inhibited the reactivation response. These results were reconciled when calcium signaling through calcineurin was shown to potentiate prostratin induced activation of NF-κB that in turn stimulated the latent HIV long terminal repeat (LTR). Similar effects of calcineurin were confirmed in a primary CD4 T cell model of HIV latency. These findings highlight an important role for calcineurin in NF-κB-dependent induction of latent HIV transcription. Innovative approaches exploiting the synergistic actions of calcineurin and prostratin in the absence of generalized T-cell activation merit exploration as a means to attack the latent viral reservoir.
Conflict of interest statement
Figures
References
-
- Prins JM, Jurriaans S, van Praag RM, Blaak H, van Rij R, et al. (1999) Immuno-activation with anti-CD3 and recombinant human IL-2 in HIV-1-infected patients on potent antiretroviral therapy. AIDS 13: 2405–2410. - PubMed
-
- Stellbrink HJ, van Lunzen J, Westby M, O’Sullivan E, Schneider C, et al. (2002) Effects of interleukin-2 plus highly active antiretroviral therapy on HIV-1 replication and proviral DNA (COSMIC trial). AIDS 16: 1479–1487. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
