

Larval defense against attack from parasitoid wasps requires nociceptive neurons
- PMID: 24205297
- PMCID: PMC3808285
- DOI: 10.1371/journal.pone.0078704
Larval defense against attack from parasitoid wasps requires nociceptive neurons
Abstract
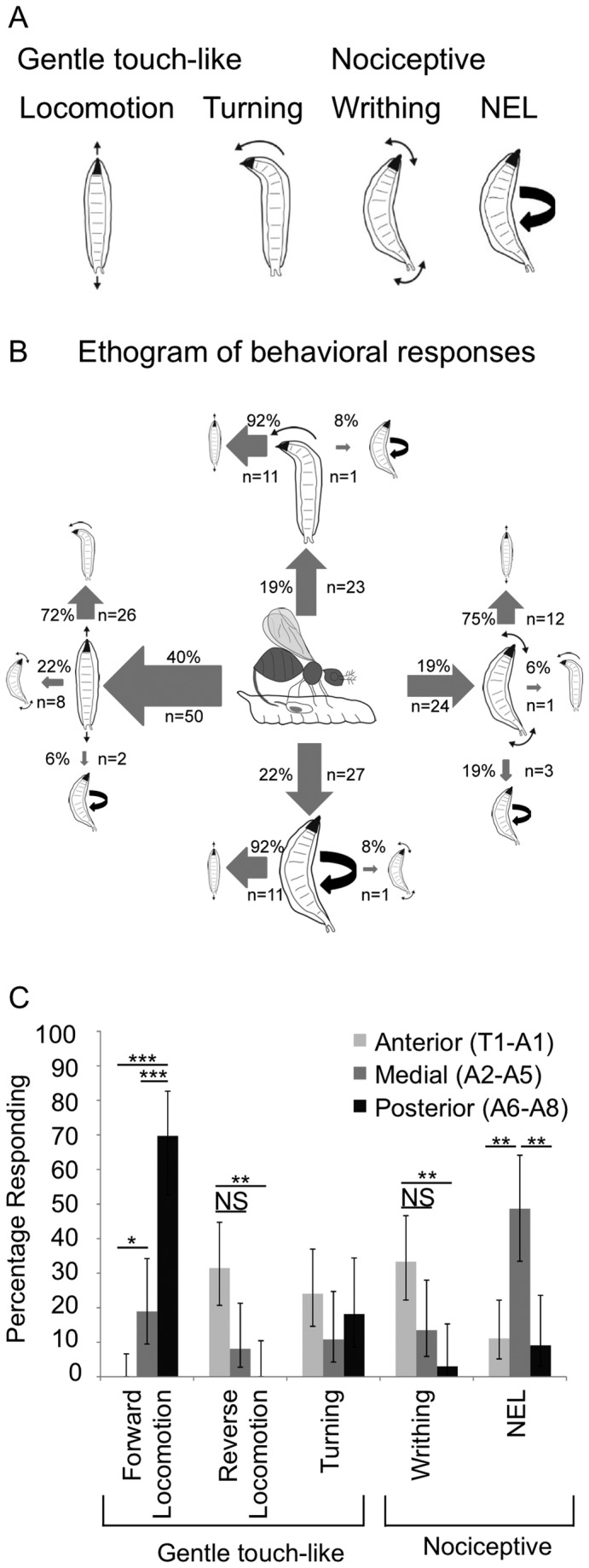
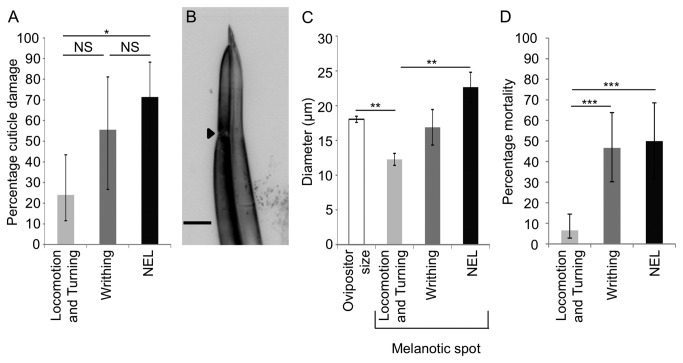
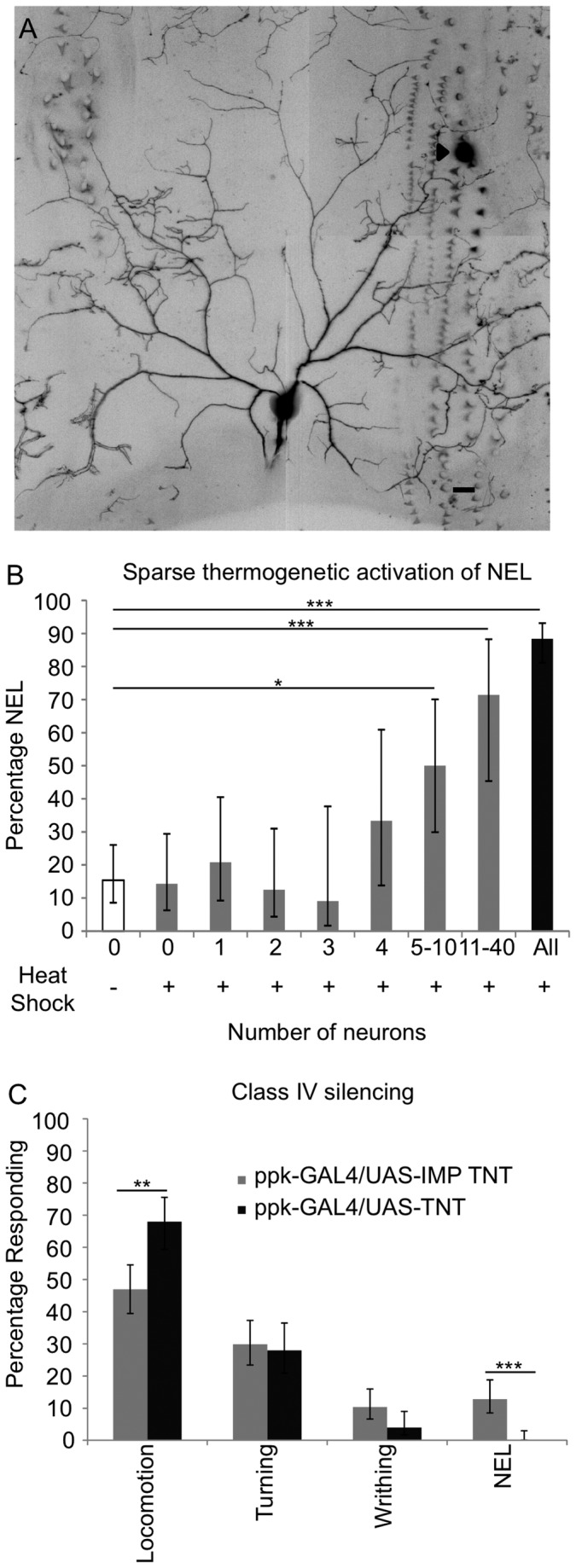
Parasitoid wasps are a fierce predator of Drosophila larvae. Female Leptopilina boulardi (LB) wasps use a sharp ovipositor to inject eggs into the bodies of Drosophila melanogaster larvae. The wasp then eats the Drosophila larva alive from the inside, and an adult wasp ecloses from the Drosophila pupal case instead of a fly. However, the Drosophila larvae are not defenseless as they may resist the attack of the wasps through somatosensory-triggered behavioral responses. Here we describe the full range of behaviors performed by the larval prey in immediate response to attacks by the wasps. Our results suggest that Drosophila larvae primarily sense the wasps using their mechanosensory systems. The range of behavioral responses included both "gentle touch" like responses as well as nociceptive responses. We found that the precise larval response depended on both the somatotopic location of the attack, and whether or not the larval cuticle was successfully penetrated during the course of the attack. Interestingly, nociceptive responses are more likely to be triggered by attacks in which the cuticle had been successfully penetrated by the wasp. Finally, we found that the class IV neurons, which are necessary for mechanical nociception, were also necessary for a nociceptive response to wasp attacks. Thus, the class IV neurons allow for a nociceptive behavioral response to a naturally occurring predator of Drosophila.
Conflict of interest statement
Figures
References
-
- Fleury F, Ris N, Allemand R, Fouillet P, Carton Y et al. (2004) Ecological and genetic interactions in Drosophila-parasitoids communities: a case study with D-melanogaster, D-simulans and their common Leptopilina parasitoids in south-eastern France. Genetica 120: 181-194. doi:10.1023/B:GENE.0000017640.78087.9e. PubMed: 15088657. - DOI - PubMed
-
- Carton Y, Bouletreau M, van Alphen JJM, van Lenteren JC (1986) The Drosophila parasitic waps. In: Ashburner M, Carson HL, Thompson JN Jr. The Genetics and Biology of Drosophila; London Academic Press. pp. 347-394
-
- Silvers MJ, Nappi AJ (1986) In vitro Study of Physiological Suppression of Supernumerary Parasites by the Endoparasitic Wasp Leptopilina Heterotoma. J Parasitol 72: 405-409. doi:10.2307/3281680. - DOI
-
- Lee MJ, Kalamarz ME, Paddibhatla I, Small C, Rajwani R et al. (2009) Virulence Factors and Strategies of Leptopilina spp.: Selective Responses in Drosophila Hosts. In: Prevost G. Advances in Parasitology, Vol 70: Parasitoids of Drosophila. San Diego: Elsevier Academic Press Inc. p. 123-+ - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
