

Electrophilic fatty acid species inhibit 5-lipoxygenase and attenuate sepsis-induced pulmonary inflammation
- PMID: 24206143
- PMCID: PMC4026401
- DOI: 10.1089/ars.2013.5473
Electrophilic fatty acid species inhibit 5-lipoxygenase and attenuate sepsis-induced pulmonary inflammation
Abstract
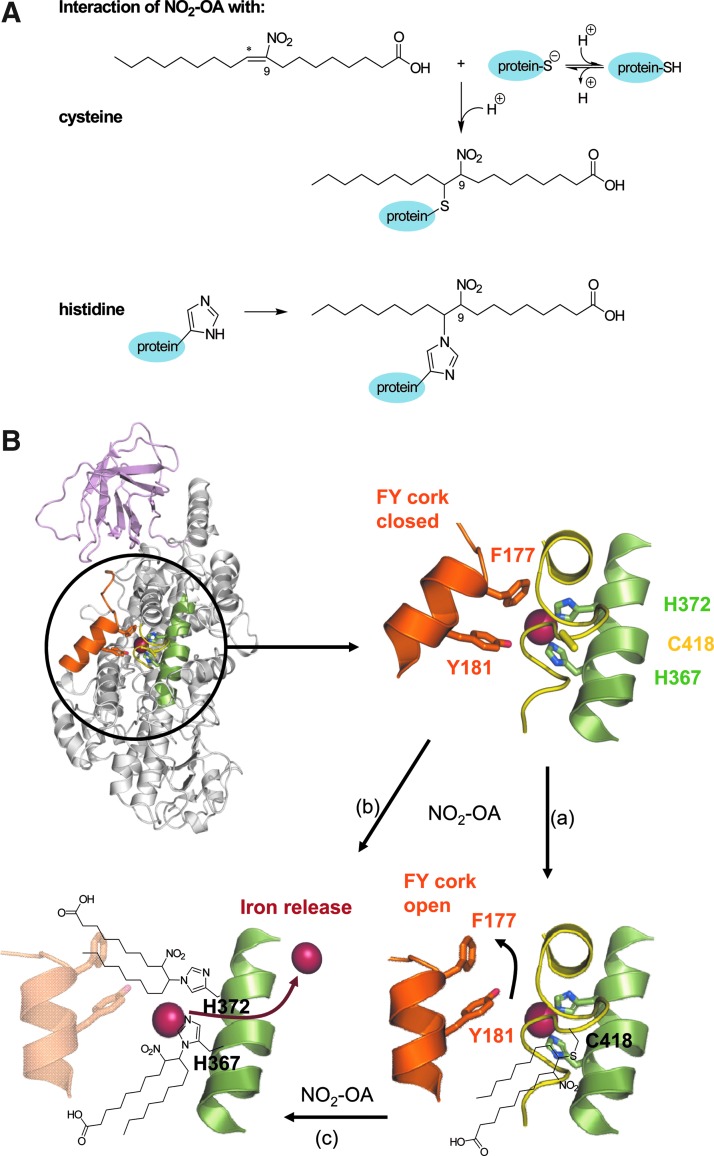
Aims: The reaction of nitric oxide and nitrite-derived species with polyunsaturated fatty acids yields electrophilic fatty acid nitroalkene derivatives (NO2-FA), which display anti-inflammatory properties. Given that the 5-lipoxygenase (5-LO, ALOX5) possesses critical nucleophilic amino acids, which are potentially sensitive to electrophilic modifications, we determined the consequences of NO2-FA on 5-LO activity in vitro and on 5-LO-mediated inflammation in vivo.
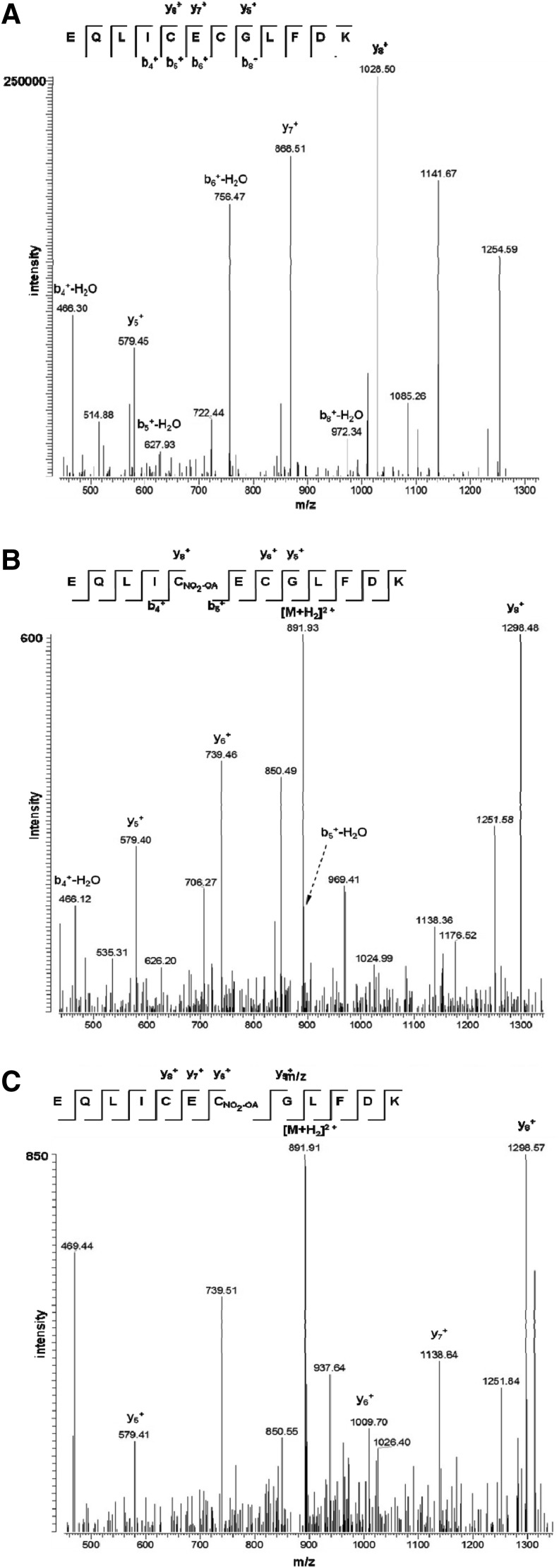
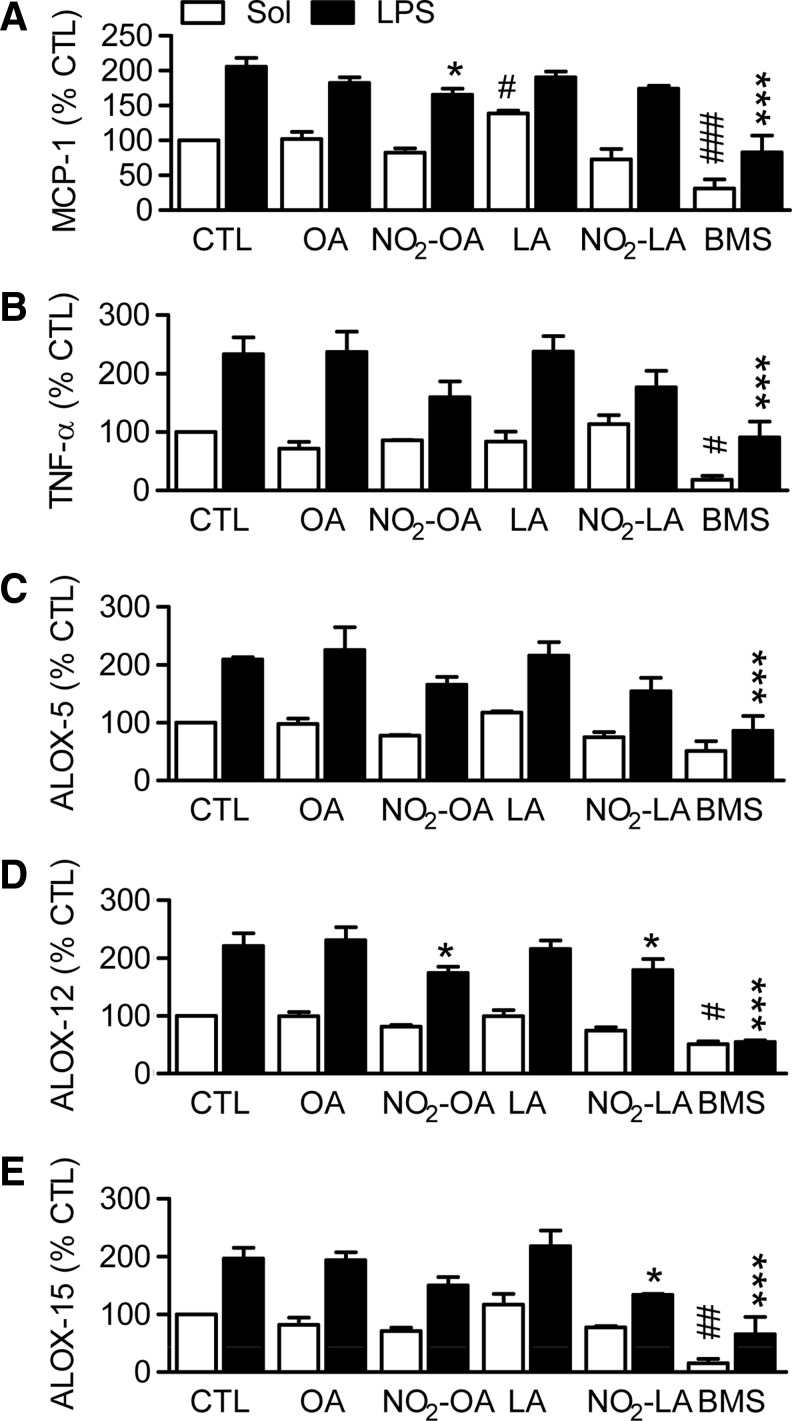
Results: Stimulation of human polymorphonuclear leukocytes (PMNL) with nitro-oleic (NO2-OA) or nitro-linoleic acid (NO2-LA) (but not the parent lipids) resulted in the concentration-dependent and irreversible inhibition of 5-LO activity. Similar effects were observed in cell lysates and using the recombinant human protein, indicating a direct reaction with 5-LO. NO2-FAs did not affect the activity of the platelet-type 12-LO (ALOX12) or 15-LO-1 (ALOX15) in intact cells or the recombinant protein. The NO2-FA-induced inhibition of 5-LO was attributed to the alkylation of Cys418, and the exchange of Cys418 to serine rendered 5-LO insensitive to NO2-FA. In vivo, the systemic administration of NO2-OA to mice decreased neutrophil and monocyte mobilization in response to lipopolysaccharide (LPS), attenuated the formation of the 5-LO product 5-hydroxyeicosatetraenoic acid (5-HETE), and inhibited lung injury. The administration of NO2-OA to 5-LO knockout mice had no effect on LPS-induced neutrophil or monocyte mobilization as well as on lung injury.
Innovation: Prophylactic administration of NO2-OA to septic mice inhibits inflammation and promotes its resolution by interfering in 5-LO-mediated inflammatory processes.
Conclusion: NO2-FAs directly and irreversibly inhibit 5-LO and attenuate downstream acute inflammatory responses.
Figures




References
-
- Baker PR, Lin Y, Schopfer FJ, Woodcock SR, Groeger AL, Batthyany C, Sweeney S, Long MH, Iles KE, Baker LM, Branchaud BP, Chen YE, and Freeman BA. Fatty acid transduction of nitric oxide signaling: multiple nitrated unsaturated fatty acid derivatives exist in human blood and urine and serve as endogenous peroxisome proliferator-activated receptor ligands. J Biol Chem 280: 42464–42475, 2005 - PMC - PubMed
-
- Ceaser EK, Moellering DR, Shiva S, Ramachandran A, Landar A, Venkartraman A, Crawford J, Patel R, Dickinson DA, Ulasova E, Ji S, and Darley-Usmar VM. Mechanisms of signal transduction mediated by oxidized lipids: the role of the electrophile-responsive proteome. Biochem Soc Trans 32: 151–155, 2004 - PubMed
-
- Chopra A, Ferreira-Alves DL, Sirois P, and Thirion JP. Cloning of the guinea pig 5-lipoxygenase gene and nucleotide sequence of its promoter. Biochem Biophys Res Commun 185: 489–495, 1992 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous