

PrimPol, an archaic primase/polymerase operating in human cells
- PMID: 24207056
- PMCID: PMC3899013
- DOI: 10.1016/j.molcel.2013.09.025
PrimPol, an archaic primase/polymerase operating in human cells
Abstract
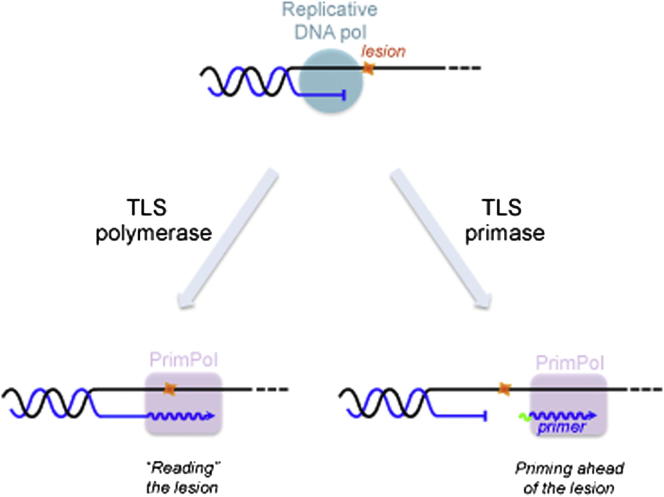
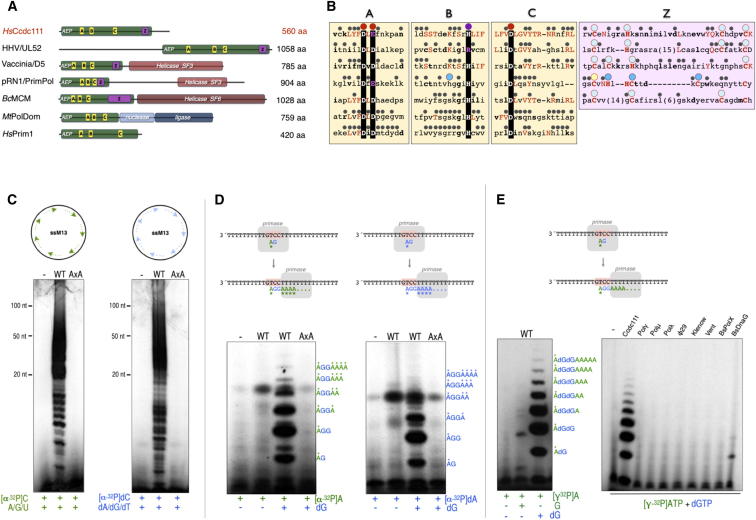
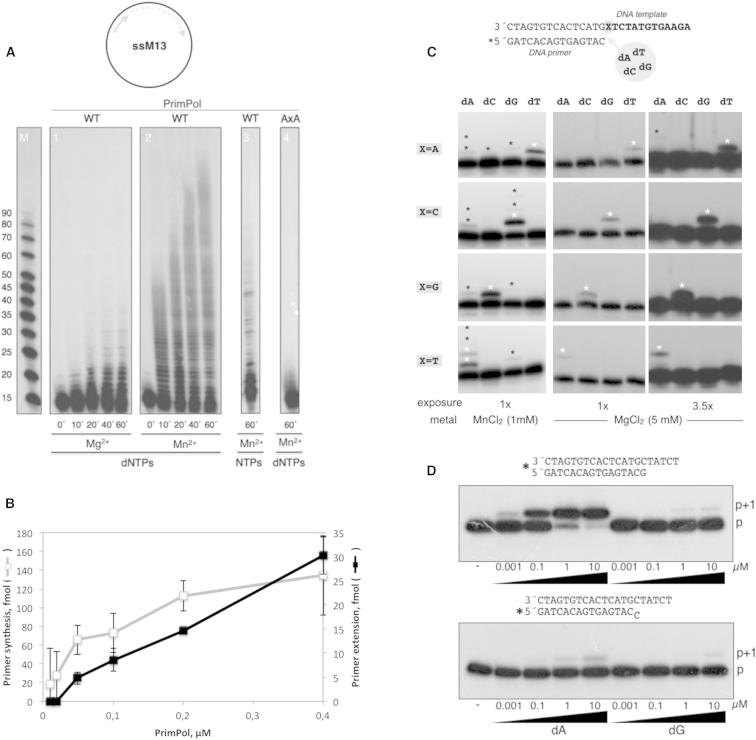
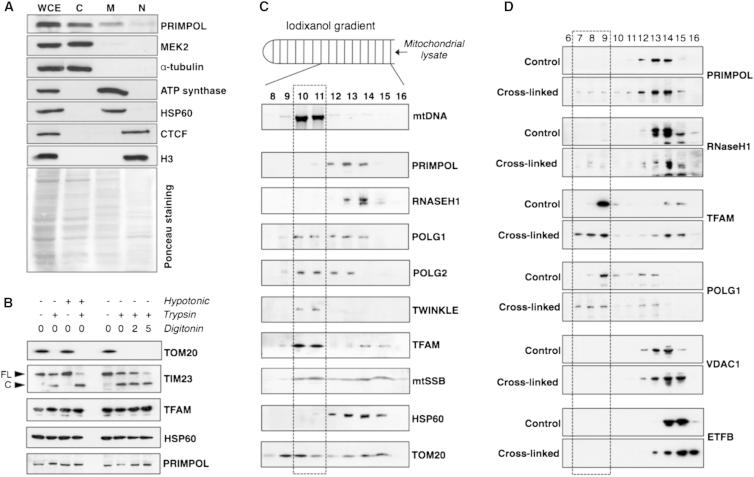
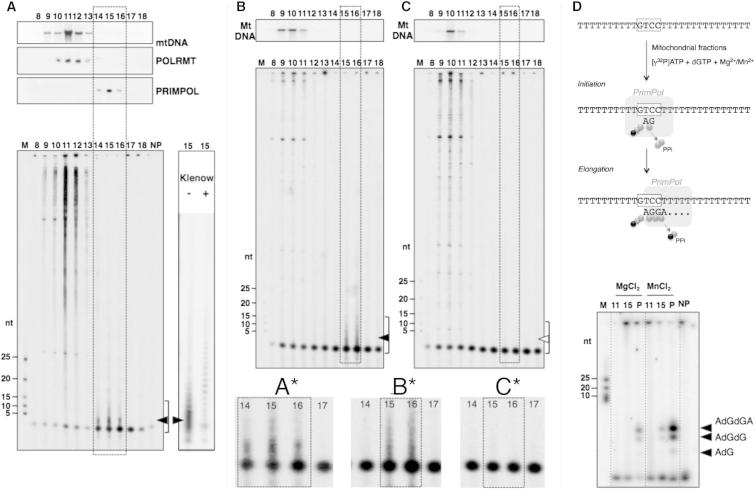
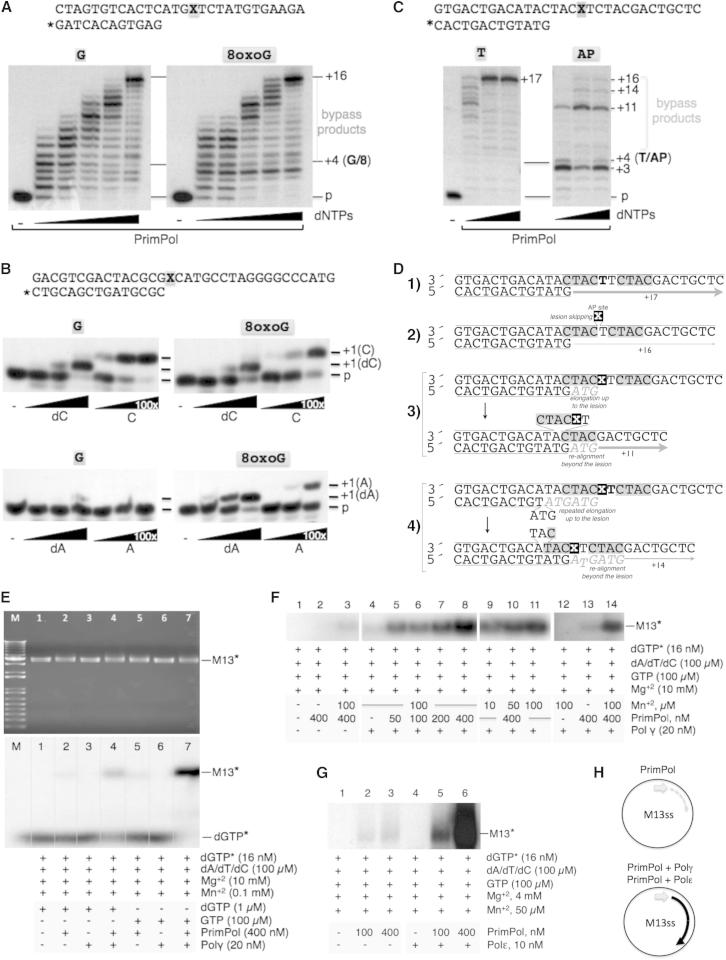
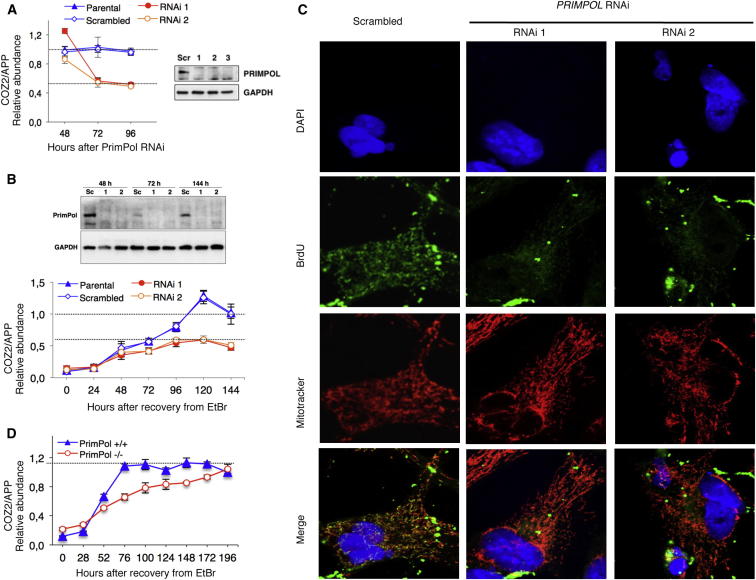
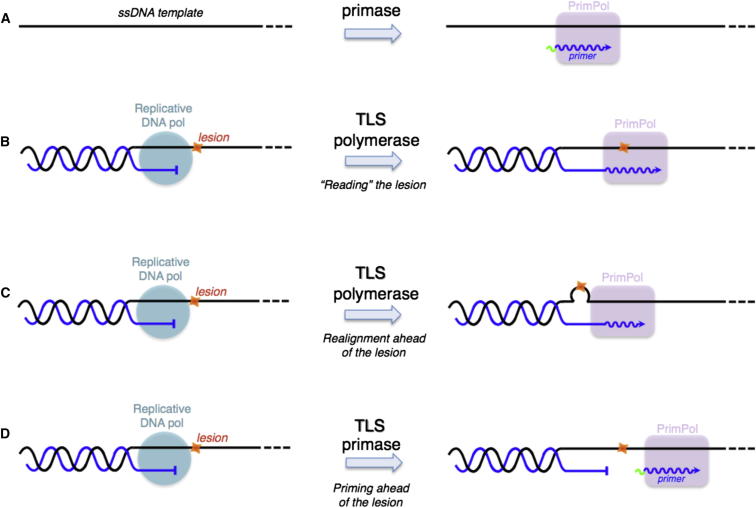
We describe a second primase in human cells, PrimPol, which has the ability to start DNA chains with deoxynucleotides unlike regular primases, which use exclusively ribonucleotides. Moreover, PrimPol is also a DNA polymerase tailored to bypass the most common oxidative lesions in DNA, such as abasic sites and 8-oxoguanine. Subcellular fractionation and immunodetection studies indicated that PrimPol is present in both nuclear and mitochondrial DNA compartments. PrimPol activity is detectable in mitochondrial lysates from human and mouse cells but is absent from mitochondria derived from PRIMPOL knockout mice. PRIMPOL gene silencing or ablation in human and mouse cells impaired mitochondrial DNA replication. On the basis of the synergy observed with replicative DNA polymerases Polγ and Polε, PrimPol is proposed to facilitate replication fork progression by acting as a translesion DNA polymerase or as a specific DNA primase reinitiating downstream of lesions that block synthesis during both mitochondrial and nuclear DNA replication.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Bernad A., Blanco L., Lázaro J.M., Martín G., Salas M. A conserved 3′----5′ exonuclease active site in prokaryotic and eukaryotic DNA polymerases. Cell. 1989;59:219–228. - PubMed
-
- Blanca G., Shevelev I., Ramadan K., Villani G., Spadari S., Hübscher U., Maga G. Human DNA polymerase lambda diverged in evolution from DNA polymerase beta toward specific Mn(++) dependence: a kinetic and thermodynamic study. Biochemistry. 2003;42:7467–7476. - PubMed
-
- Bolden A., Noy G.P., Weissbach A. DNA polymerase of mitochondria is a gamma-polymerase. J. Biol. Chem. 1977;252:3351–3356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
