

A beneficial role for immunoglobulin E in host defense against honeybee venom
- PMID: 24210352
- PMCID: PMC4164235
- DOI: 10.1016/j.immuni.2013.10.005
A beneficial role for immunoglobulin E in host defense against honeybee venom
Abstract
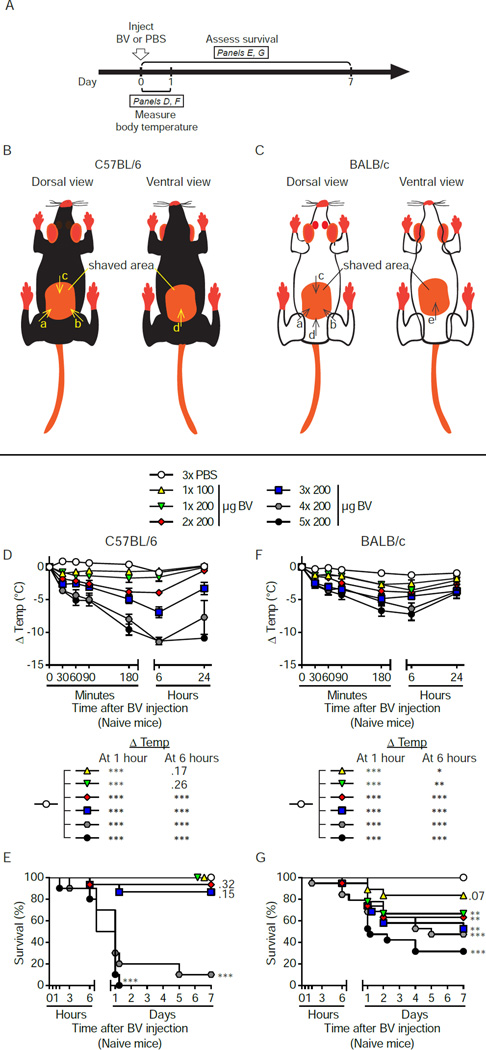
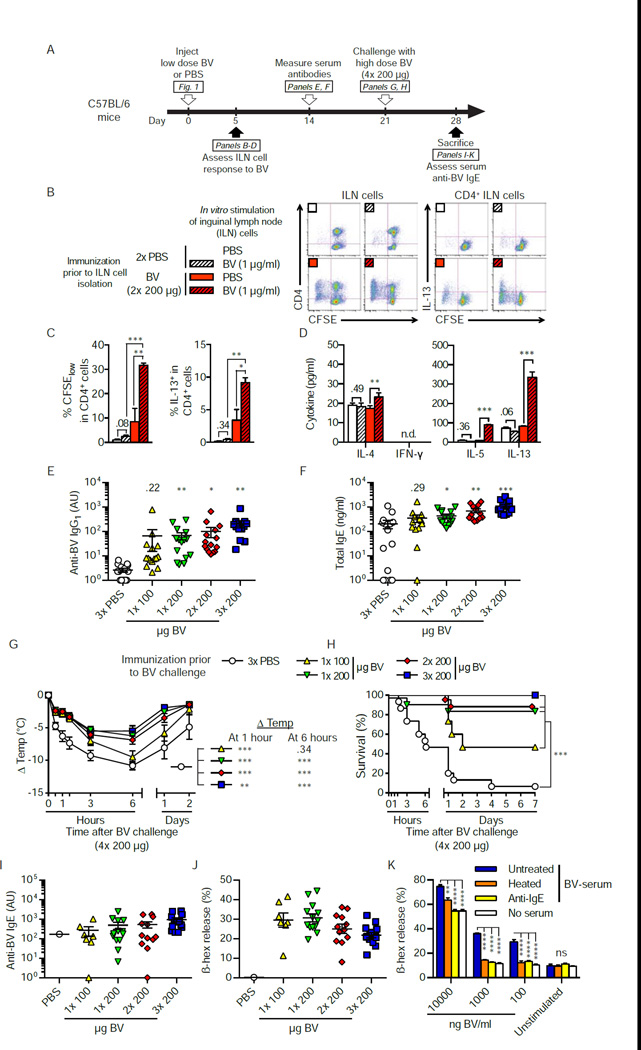
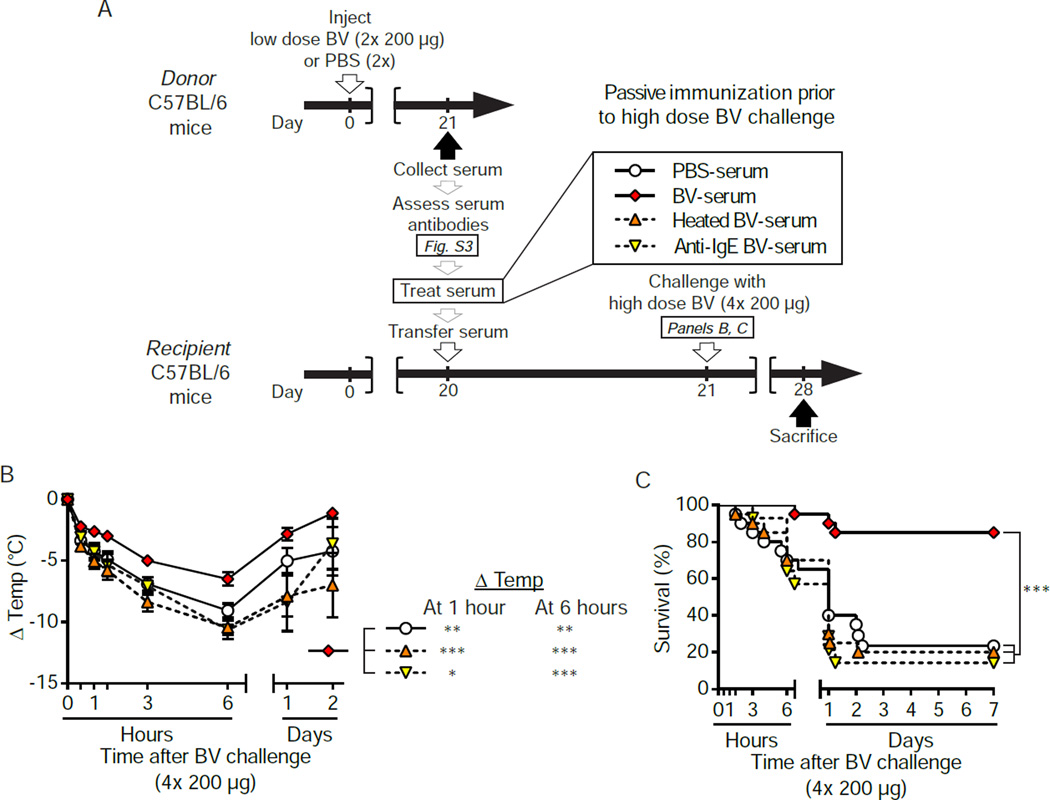
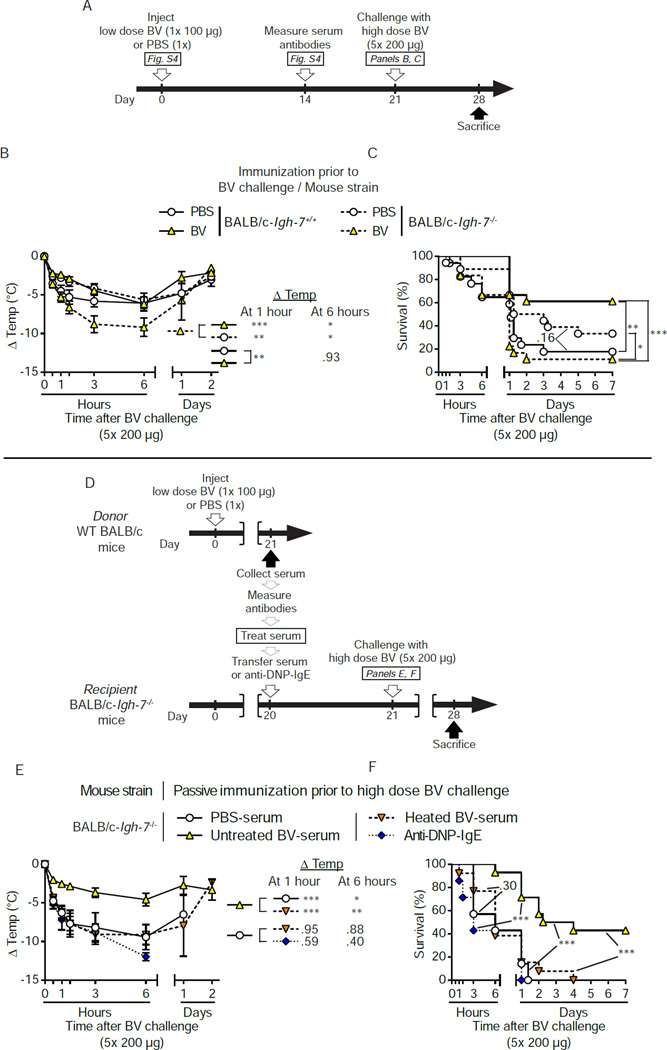
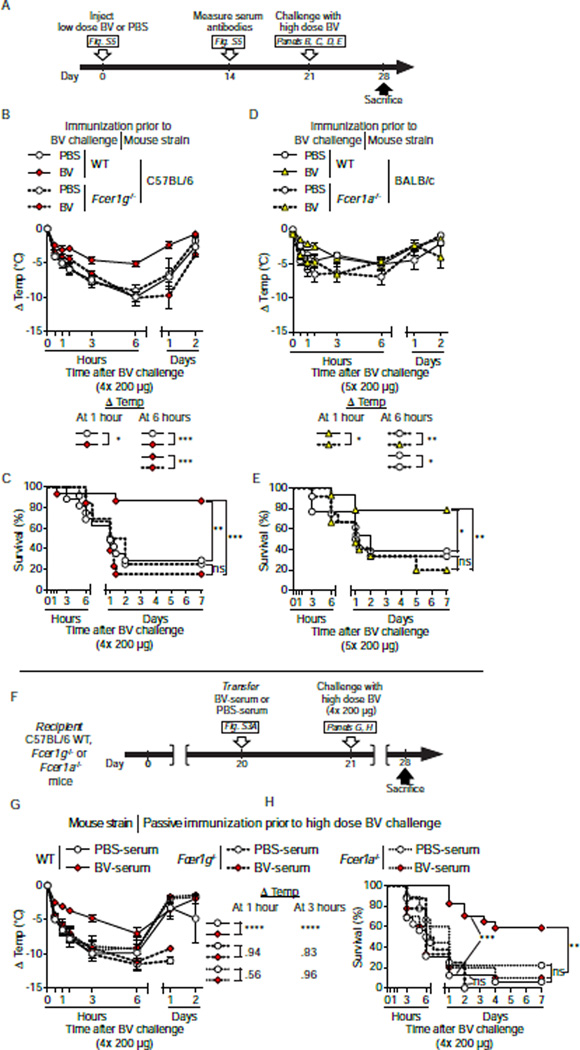
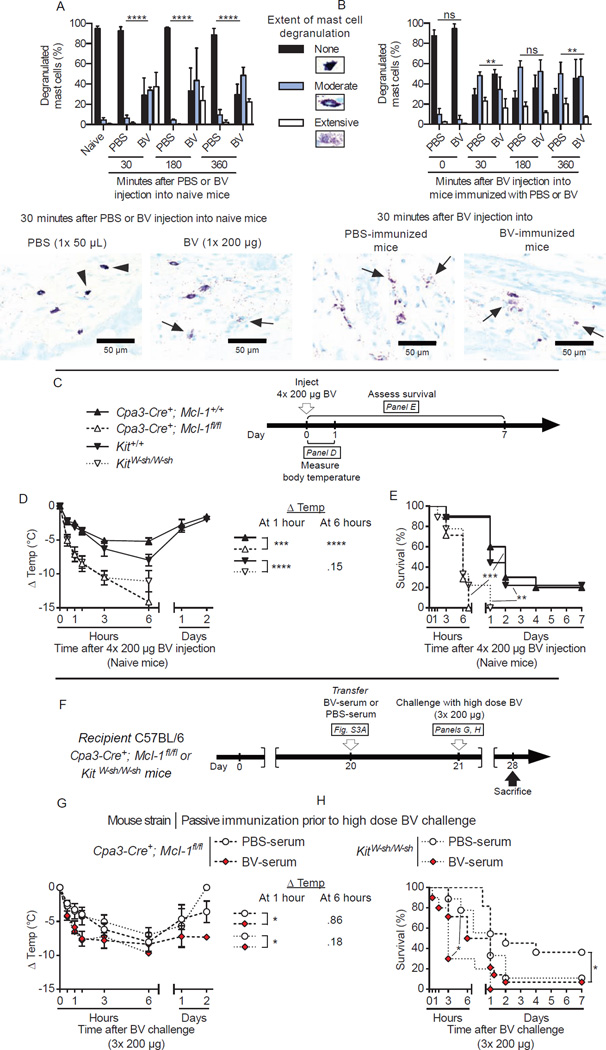
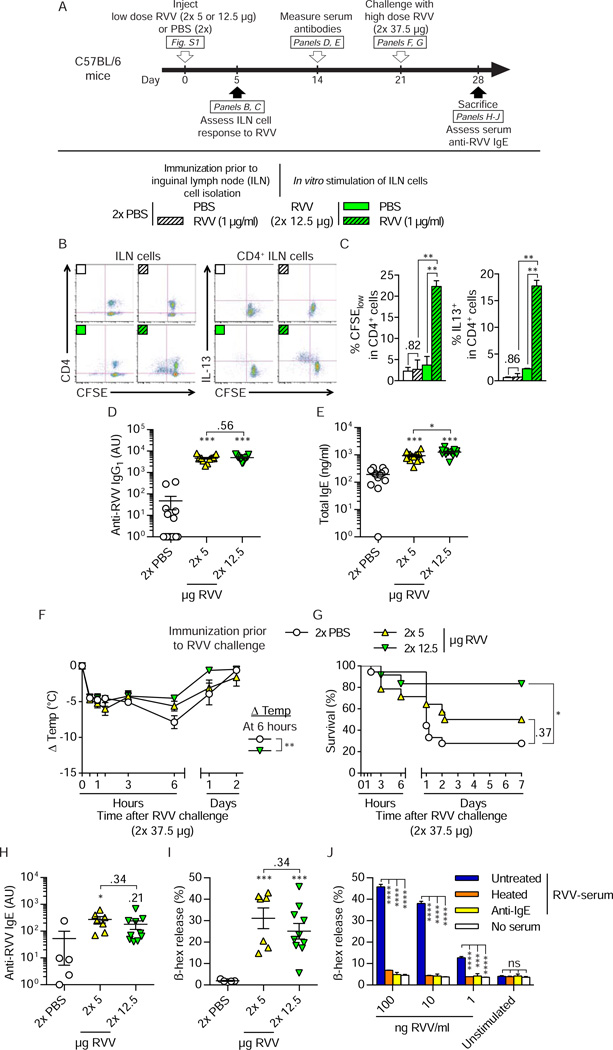
Allergies are widely considered to be misdirected type 2 immune responses, in which immunoglobulin E (IgE) antibodies are produced against any of a broad range of seemingly harmless antigens. However, components of insect venoms also can sensitize individuals to develop severe IgE-associated allergic reactions, including fatal anaphylaxis, upon subsequent venom exposure. We found that mice injected with amounts of honeybee venom similar to that which could be delivered in one or two stings developed a specific type 2 immune response that increased their resistance to subsequent challenge with potentially lethal amounts of the venom. Our data indicate that IgE antibodies and the high affinity IgE receptor, FcεRI, were essential for such acquired resistance to honeybee venom. The evidence that IgE-dependent immune responses against venom can enhance survival in mice supports the hypothesis that IgE, which also contributes to allergic disorders, has an important function in protection of the host against noxious substances.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Antibodies: taking the sting out.Nat Rev Immunol. 2013 Dec;13(12):843. doi: 10.1038/nri3568. Epub 2013 Nov 8. Nat Rev Immunol. 2013. PMID: 24202657 No abstract available.
-
A sting in the tale of TH2 immunity.Immunity. 2013 Nov 14;39(5):803-5. doi: 10.1016/j.immuni.2013.10.015. Immunity. 2013. PMID: 24238337
References
-
- Akahoshi M, Song CH, Piliponsky AM, Metz M, Guzzetta A, Abrink M, Schlenner SM, Feyerabend TB, Rodewald HR, Pejler G, et al. Mast cell chymase reduces the toxicity of Gila monster venom, scorpion venom, and vasoactive intestinal polypeptide in mice. J. Clin. Invest. 2011;121:4180–4191. - PMC - PubMed
-
- Ando A, Martin TR, Galli SJ. Effects of chronic treatment with the c-kit ligand, stem cell factor, on immunoglobulin E-dependent anaphylaxis in mice. Genetically mast cell-deficient Sl/Sld mice acquire anaphylactic responsiveness, but the congenic normal mice do not exhibit augmented responses. J. Clin. Invest. 1993;92:1639–1649. - PMC - PubMed
-
- Annila I. Bee venom allergy. Clin. Exp. Allergy. 2000;30:1682–1687. - PubMed
-
- Antonicelli L, Bilo MB, Bonifazi F. Epidemiology of Hymenoptera allergy. Curr. Opin. Allergy Clin. Immunol. 2002;2:341–346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
