

SHARPIN regulates uropod detachment in migrating lymphocytes
- PMID: 24210817
- PMCID: PMC3852511
- DOI: 10.1016/j.celrep.2013.10.011
SHARPIN regulates uropod detachment in migrating lymphocytes
Abstract
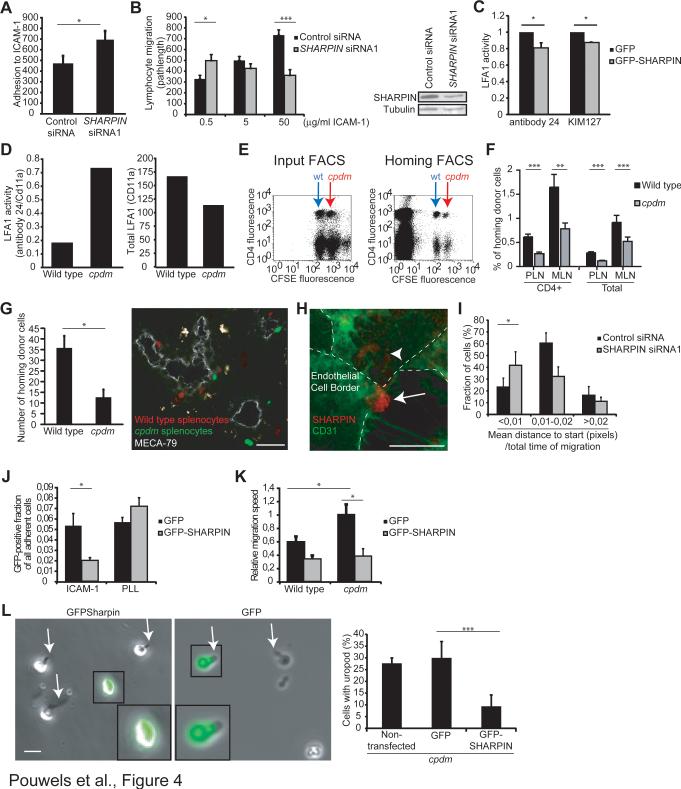
SHARPIN-deficient mice display a multiorgan chronic inflammatory phenotype suggestive of altered leukocyte migration. We therefore studied the role of SHARPIN in lymphocyte adhesion, polarization, and migration. We found that SHARPIN localizes to the trailing edges (uropods) of both mouse and human chemokine-activated lymphocytes migrating on intercellular adhesion molecule-1 (ICAM-1), which is one of the major endothelial ligands for migrating leukocytes. SHARPIN-deficient cells adhere better to ICAM-1 and show highly elongated tails when migrating. The increased tail lifetime in SHARPIN-deficient lymphocytes decreases the migration velocity. The adhesion, migration, and uropod defects in SHARPIN-deficient lymphocytes were rescued by reintroducing SHARPIN into the cells. Mechanistically, we show that SHARPIN interacts directly with lymphocyte-function-associated antigen-1 (LFA-1), a leukocyte counterreceptor for ICAM-1, and inhibits the expression of intermediate and high-affinity forms of LFA-1. Thus, SHARPIN controls lymphocyte migration by endogenously maintaining LFA-1 inactive to allow adjustable detachment of the uropods in polarized cells.
Copyright © 2013 The Authors. Published by Elsevier Inc. All rights reserved.
Figures



References
-
- Bolomini-Vittori M, Montresor A, Giagulli C, Staunton D, Rossi B, Martinello M, Constantin G, Laudanna C. Regulation of conformer-specific activation of the integrin LFA-1 by a chemokine-triggered Rho signaling module. Nat Immunol. 2009;10:185–194. - PubMed
-
- Constantin G, Majeed M, Giagulli C, Piccio L, Kim JY, Butcher EC, Laudanna C. Chemokines trigger immediate beta2 integrin affinity and mobility changes: differential regulation and roles in lymphocyte arrest under flow. Immunity. 2000;13:759–769. - PubMed
-
- Franco SJ, Rodgers MA, Perrin BJ, Han J, Bennin DA, Critchley DR, Huttenlocher A. Calpain-mediated proteolysis of talin regulates adhesion dynamics. Nat Cell Biol. 2004;6:977–983. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
