

Changes in cellular mRNA stability, splicing, and polyadenylation through HuR protein sequestration by a cytoplasmic RNA virus
- PMID: 24210824
- PMCID: PMC3849337
- DOI: 10.1016/j.celrep.2013.10.012
Changes in cellular mRNA stability, splicing, and polyadenylation through HuR protein sequestration by a cytoplasmic RNA virus
Abstract
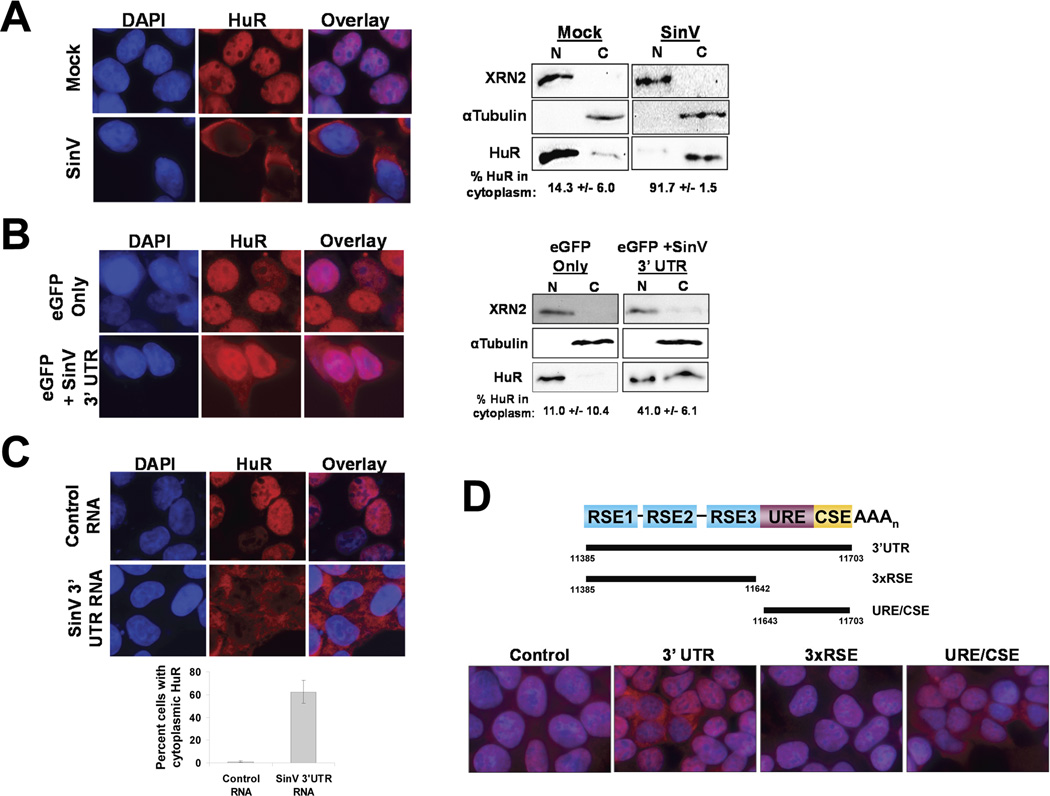
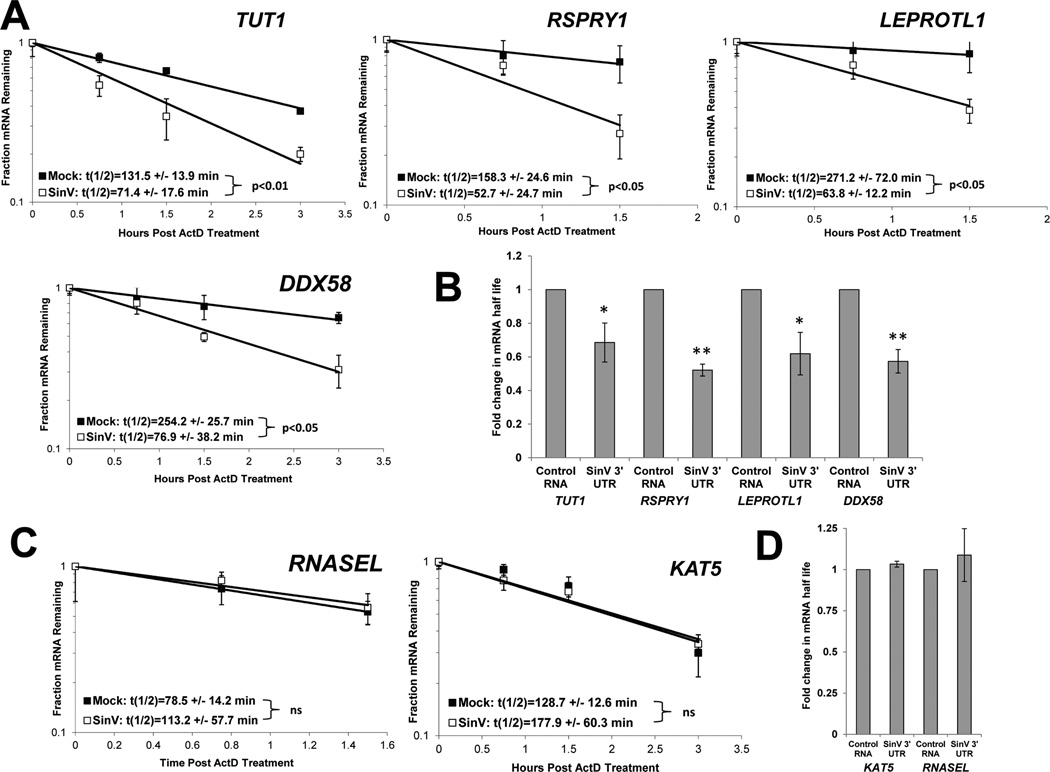
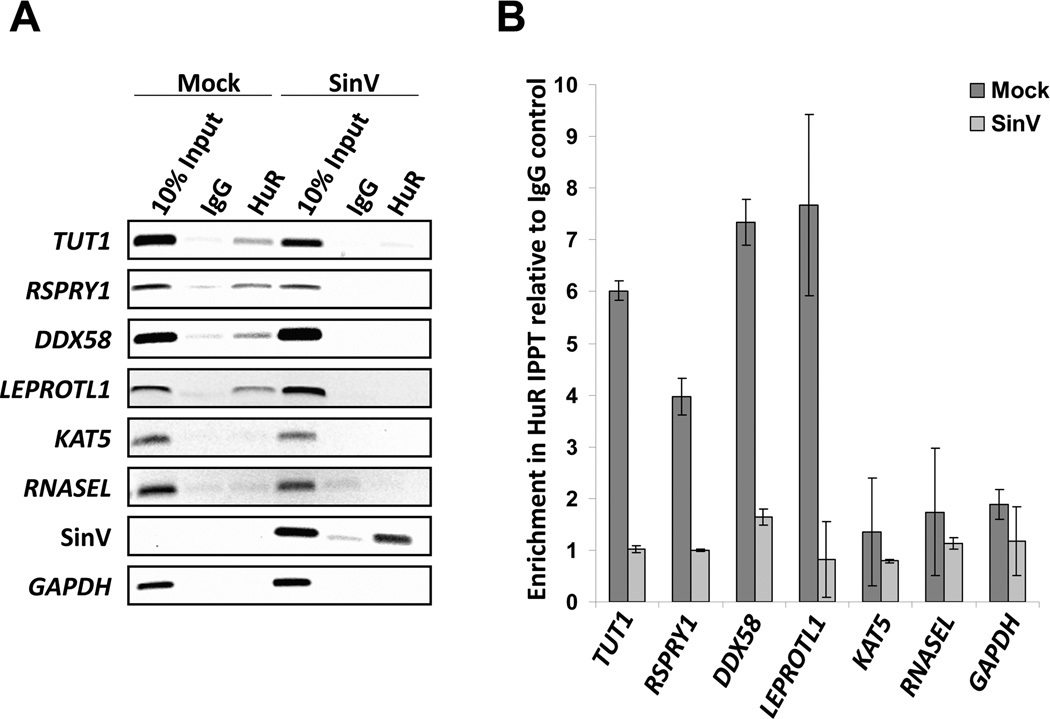
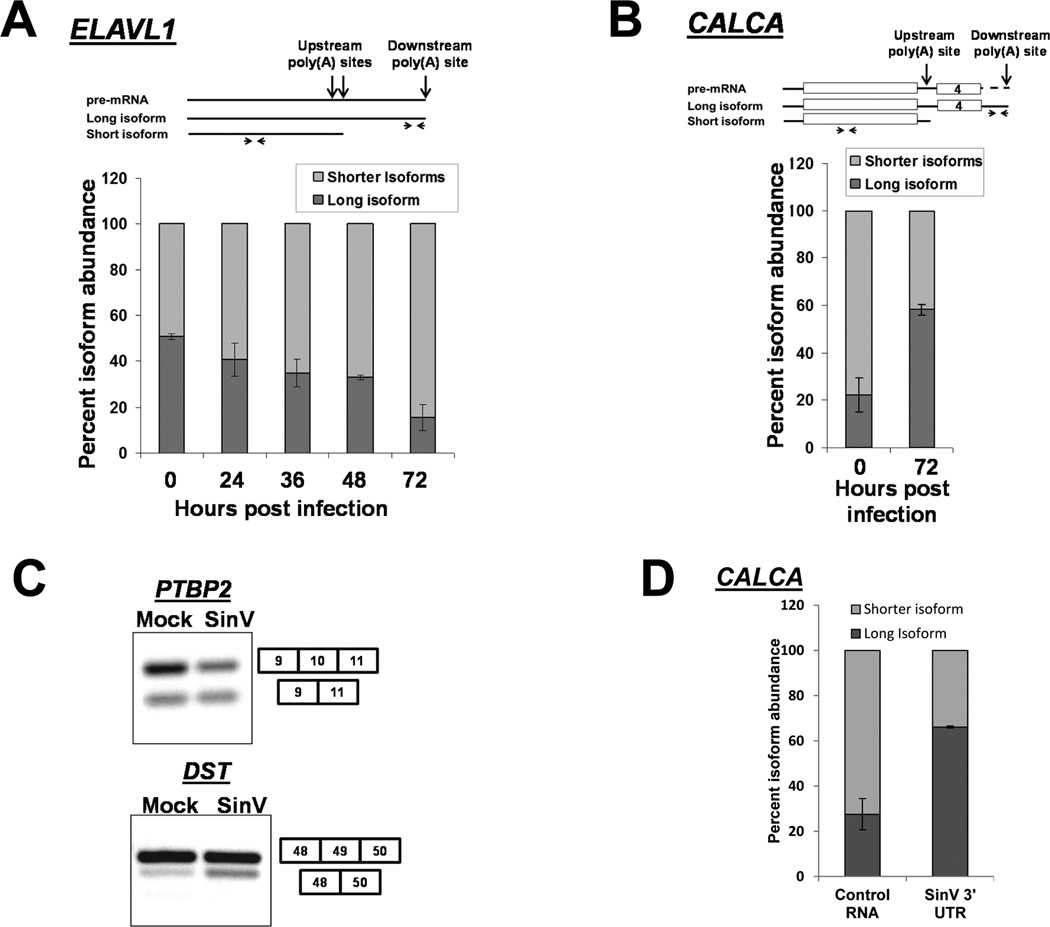
The impact of RNA viruses on the posttranscriptional regulation of cellular gene expression is unclear. Sindbis virus causes a dramatic relocalization of the cellular HuR protein from the nucleus to the cytoplasm in infected cells. This is to the result of the expression of large amounts of viral RNAs that contain high-affinity HuR binding sites in their 3' UTRs effectively serving as a sponge for the HuR protein. Sequestration of HuR by Sindbis virus is associated with destabilization of cellular mRNAs that normally bind HuR and rely on it to regulate their expression. Furthermore, significant changes can be observed in nuclear alternative polyadenylation and splicing events on cellular pre-mRNAs as a result of sequestration of HuR protein by the 3' UTR of transcripts of this cytoplasmic RNA virus. These studies suggest a molecular mechanism of virus-host interaction that probably has a significant impact on virus replication, cytopathology, and pathogenesis.
Copyright © 2013 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Benoit RM, Meisner NC, Kallen J, Graff P, Hemmig R, Cèbe R, Ostermeier C, Widmer H, Auer M. The x-ray crystal structure of the first RNA recognition motif and site-directed mutagenesis suggest a possible HuR redox sensing mechanism. J. Mol. Biol. 2010;397:1231–1244. - PubMed
-
- Bevilacqua A, Ghisolfi L, Franzi S, Maresca G, Gherzi R, Capaccioli S, Nicolin A, Canti G. Stabilization of cellular mRNAs and up-regulation of proteins by oligoribonucleotides homologous to the Bcl2 adenine-uridine rich element motif. Mol. Pharmacol. 2007;71:531–538. - PubMed
-
- Bolognani F, Gallani AI, Sokol L, Baskin DS, Meisner-Kober N. mRNA stability alterations mediated by HuR are necessary to sustain the fast growth of glioma cells. J. Neurooncol. 2012;106:531–542. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
