

Type IV pilus biogenesis, twitching motility, and DNA uptake in Thermus thermophilus: discrete roles of antagonistic ATPases PilF, PilT1, and PilT2
- PMID: 24212586
- PMCID: PMC3911100
- DOI: 10.1128/AEM.03218-13
Type IV pilus biogenesis, twitching motility, and DNA uptake in Thermus thermophilus: discrete roles of antagonistic ATPases PilF, PilT1, and PilT2
Abstract
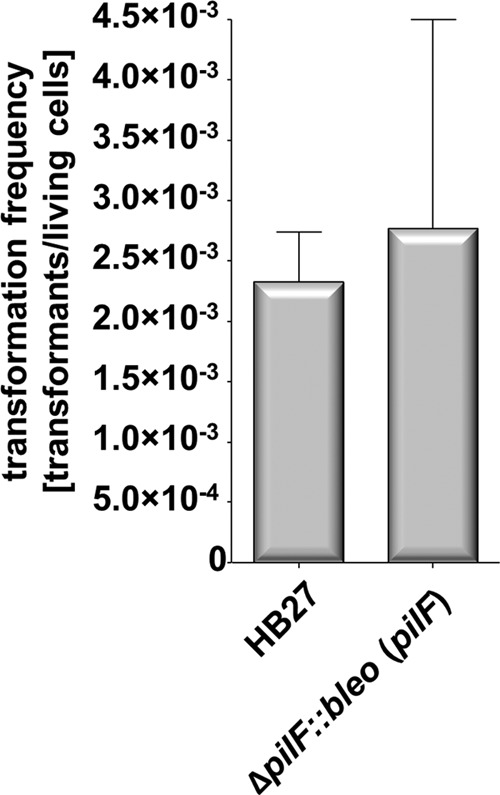
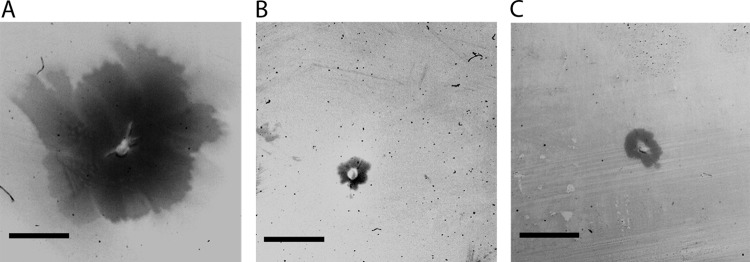
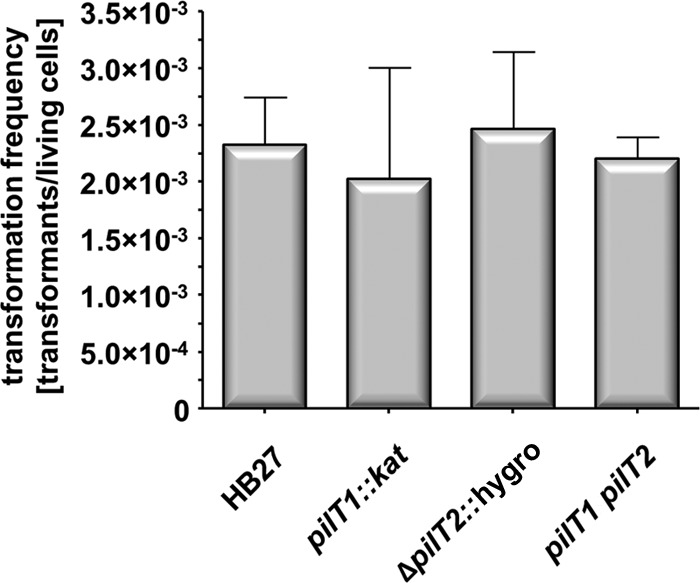
Natural transformation has a large impact on lateral gene flow and has contributed significantly to the ecological diversification and adaptation of bacterial species. Thermus thermophilus HB27 has emerged as the leading model organism for studies of DNA transporters in thermophilic bacteria. Recently, we identified a zinc-binding polymerization nucleoside triphosphatase (NTPase), PilF, which is essential for the transport of DNA through the outer membrane. Here, we present genetic evidence that PilF is also essential for the biogenesis of pili. One of the most challenging questions was whether T. thermophilus has any depolymerization NTPase acting as a counterplayer of PilF. We identified two depolymerization NTPases, PilT1 (TTC1621) and PilT2 (TTC1415), both of which are required for type IV pilus (T4P)-mediated twitching motility and adhesion but dispensable for natural transformation. This suggests that T4P dynamics are not required for natural transformation. The latter finding is consistent with our suggestion that in T. thermophilus, T4P and natural transformation are linked but distinct systems.
Figures
References
-
- Hobbs M, Mattick JS. 1993. Common components in the assembly of type 4 fimbriae, DNA transfer systems, filamentous phage and protein-secretion apparatus: a general system for the formation of surface-associated protein complexes. Mol. Microbiol. 10:233–243. 10.1111/j.1365-2958.1993.tb01949.x - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
