

A chain mechanism for flagellum growth
- PMID: 24213633
- PMCID: PMC3864836
- DOI: 10.1038/nature12682
A chain mechanism for flagellum growth
Abstract
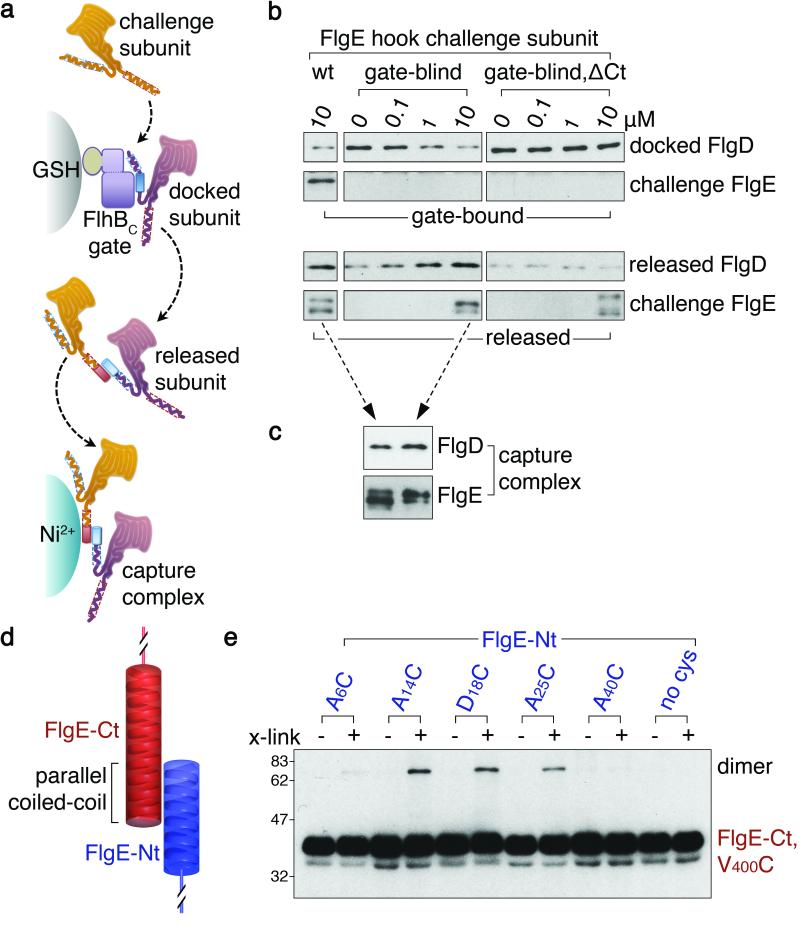
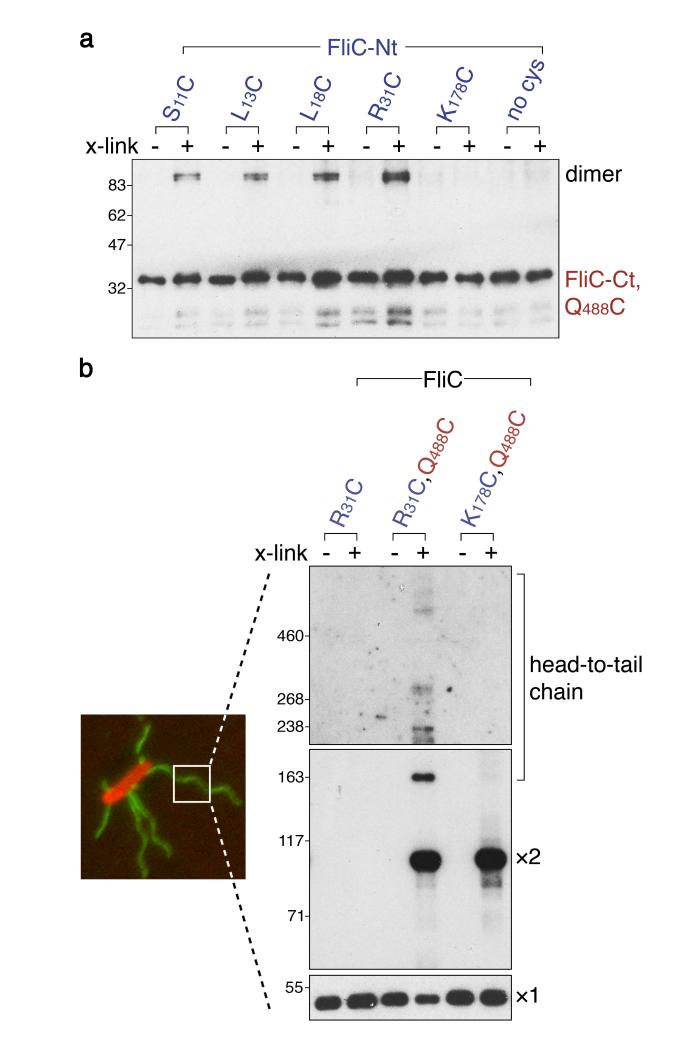
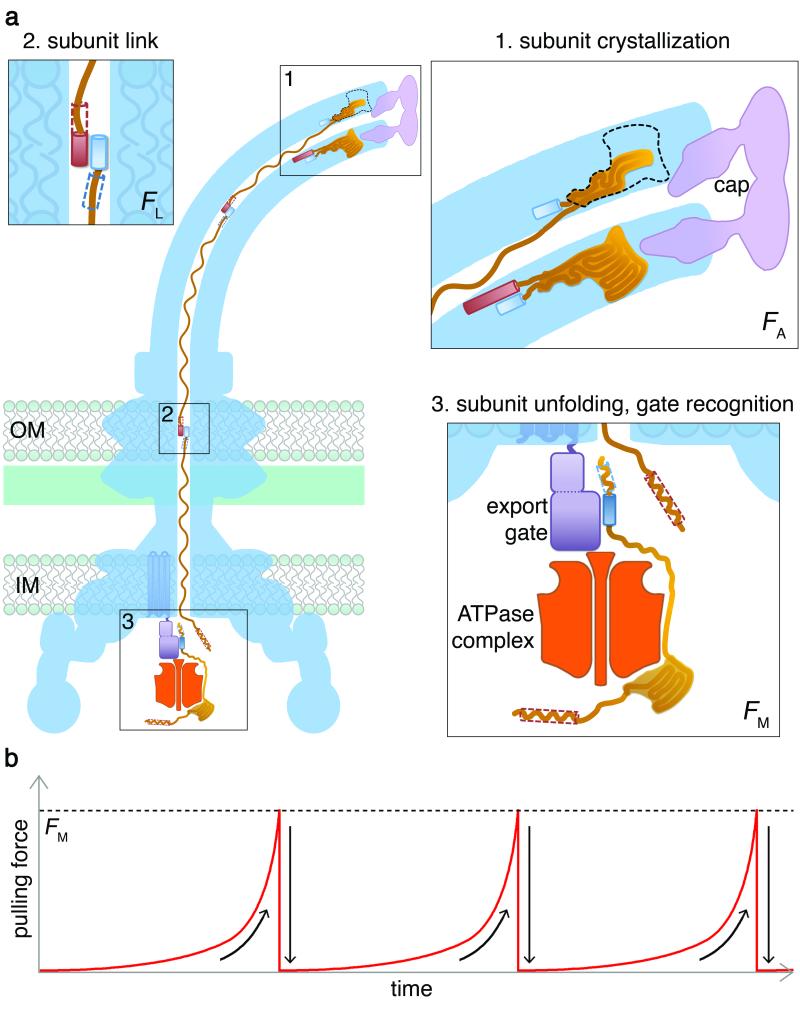
Bacteria swim by means of long flagella extending from the cell surface. These are assembled from thousands of protein subunits translocated across the cell membrane by an export machinery at the base of each flagellum. Unfolded subunits then transit through a narrow channel at the core of the growing flagellum to the tip, where they crystallize into the nascent structure. As the flagellum lengthens outside the cell, the rate of flagellum growth does not change. The mystery is how subunit transit is maintained at a constant rate without a discernible energy source in the channel of the external flagellum. We present evidence for a simple physical mechanism for flagellum growth that harnesses the entropic force of the unfolded subunits themselves. We show that a subunit docked at the export machinery can be captured by a free subunit through head-to-tail linkage of juxtaposed amino (N)- and carboxy (C)-terminal helices. We propose that sequential rounds of linkage would generate a multisubunit chain that pulls successive subunits into and through the channel to the flagellum tip, and by isolating filaments growing on bacterial cells we reveal the predicted chain of head-to-tail linked subunits in the transit channel of flagella. Thermodynamic analysis confirms that links in the subunit chain can withstand the pulling force generated by rounds of subunit crystallization at the flagellum tip, and polymer theory predicts that as the N terminus of each unfolded subunit crystallizes, the entropic force at the subunit C terminus would increase, rapidly overcoming the threshold required to pull the next subunit from the export machinery. This pulling force would adjust automatically over the increasing length of the growing flagellum, maintaining a constant rate of subunit delivery to the tip.
Figures
References
-
- Akeda Y, Galán JE. Chaperone release and unfolding of substrates in type III secretion. Nature. 2005;437:911–915. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
