

Implicating SCF complexes in organogenesis in Caenorhabditis elegans
- PMID: 24214340
- PMCID: PMC3872186
- DOI: 10.1534/genetics.113.158485
Implicating SCF complexes in organogenesis in Caenorhabditis elegans
Abstract
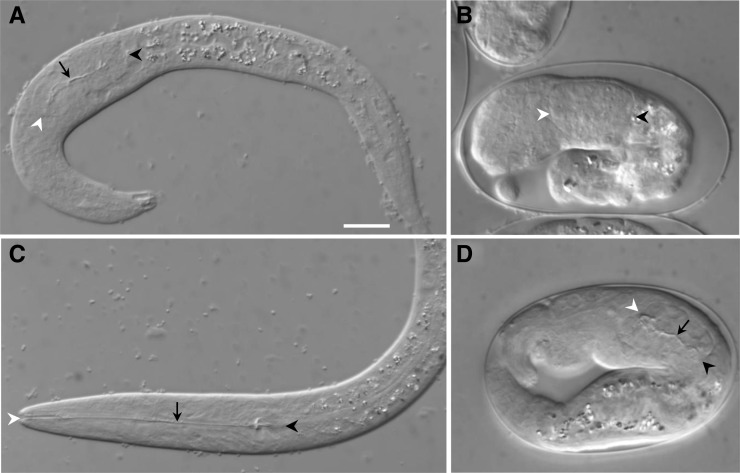
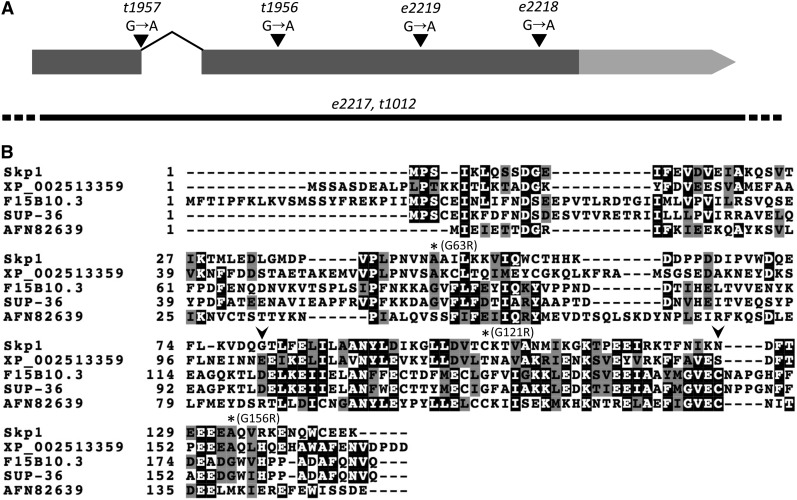
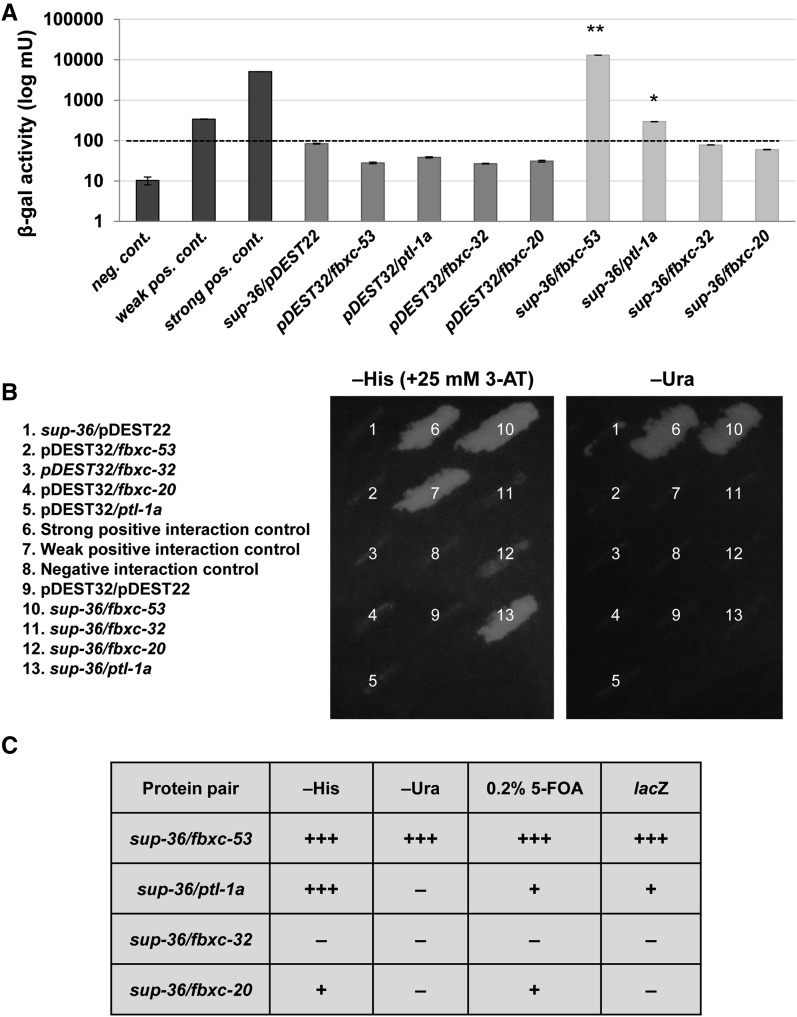
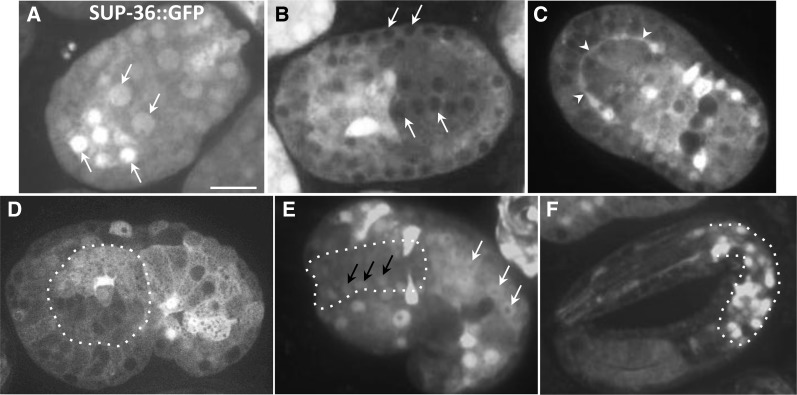
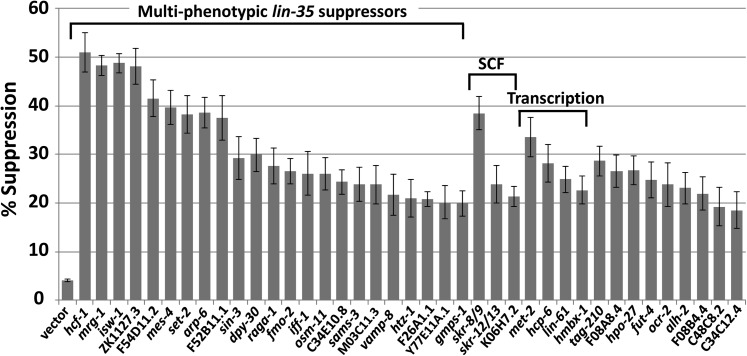
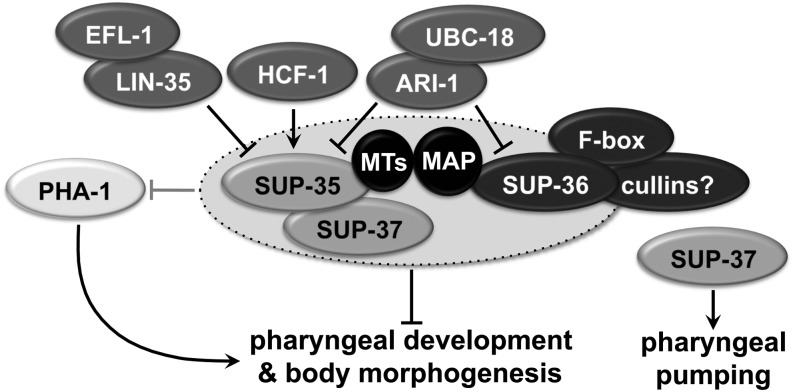
Development of the Caenorhabditis elegans foregut (pharynx) is regulated by a network of proteins that includes the Retinoblastoma protein (pRb) ortholog LIN-35; the ubiquitin pathway components UBC-18 and ARI-1; and PHA-1, a cytoplasmic protein. Loss of pha-1 activity impairs pharyngeal development and body morphogenesis, leading to embryonic arrest. We have used a genetic suppressor approach to dissect this complex pathway. The lethality of pha-1 mutants is suppressed by loss-of-function mutations in sup-35/ztf-21 and sup-37/ztf-12, which encode Zn-finger proteins, and by mutations in sup-36. Here we show that sup-36 encodes a divergent Skp1 family member that binds to several F-box proteins and the microtubule-associated protein PLT-1/τ. Like SUP-35, SUP-36 levels were negatively regulated by UBC-18-ARI-1. We also found that SUP-35 and SUP-37 physically associated and that SUP-35 could bind microtubules. Thus, SUP-35, SUP-36, and SUP-37 may function within a pathway or complex that includes cytoskeletal components. Additionally, SUP-36 may regulate the subcellular localization of SUP-35 during embryogenesis. We carried out a genome-wide RNAi screen to identify additional regulators of this network and identified 39 genes, most of which are associated with transcriptional regulation. Twenty-three of these genes acted via the LIN-35 pathway. In addition, several S-phase kinase-associated protein (Skp)1-Cullin-F-Box (SCF) components were identified, further implicating SCF complexes as part of the greater network controlling pharyngeal development.
Keywords: Caenorhabditis elegans; F-box; Skp1 pharynx; ari-1; lin-35; morphogenesis; pha-1; ptl-1; sup-35; sup-36; sup-37; ubc-18.
Figures
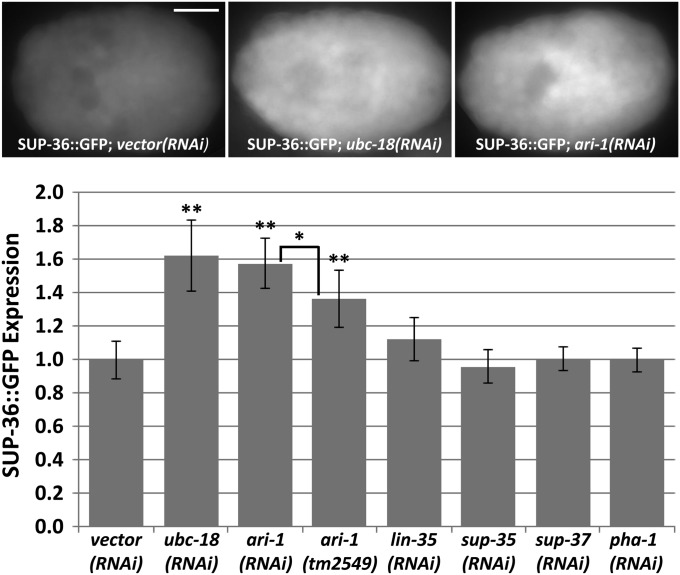
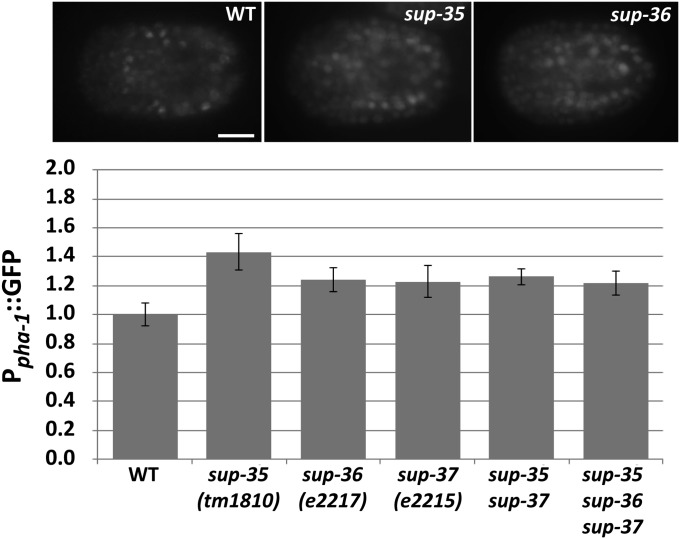
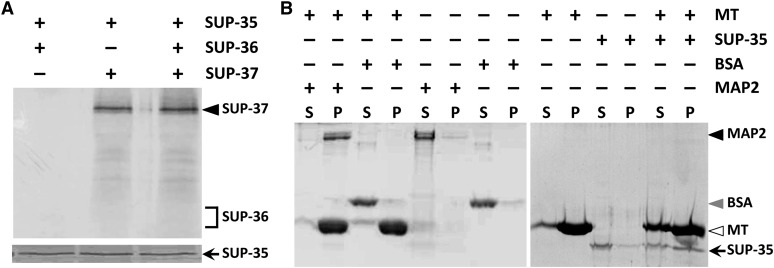
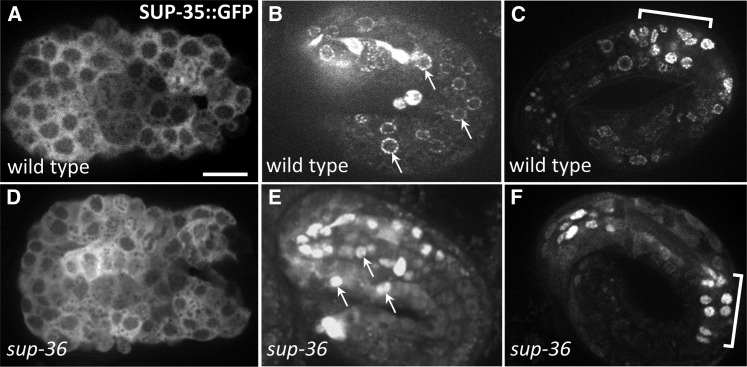
References
-
- Ahringer, J., 2005 Reverse genetics (April 6, 2006), WormBook, ed. The C. elegans Research Community, WormBook, /10.1895/wormbook.1.47.1, http://www.wormbook.org.
-
- Andersen E. C., Horvitz H. R., 2007. Two C. elegans histone methyltransferases repress lin-3 EGF transcription to inhibit vulval development. Development 134: 2991–2999. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
