

RBF1, a plant homolog of the bacterial ribosome-binding factor RbfA, acts in processing of the chloroplast 16S ribosomal RNA
- PMID: 24214533
- PMCID: PMC3875801
- DOI: 10.1104/pp.113.228338
RBF1, a plant homolog of the bacterial ribosome-binding factor RbfA, acts in processing of the chloroplast 16S ribosomal RNA
Erratum in
-
CORRECTIONS.Plant Physiol. 2015 Oct;169(2):1419. doi: 10.1104/pp.15.01381. Plant Physiol. 2015. PMID: 26417055 Free PMC article. No abstract available.
Abstract
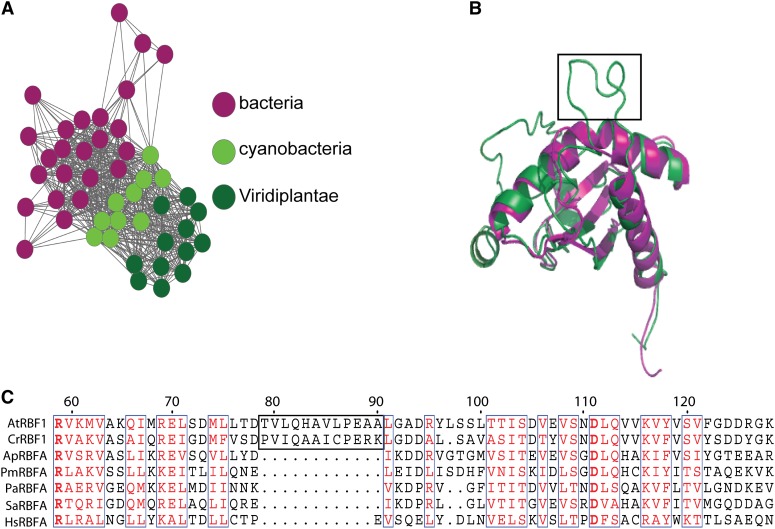
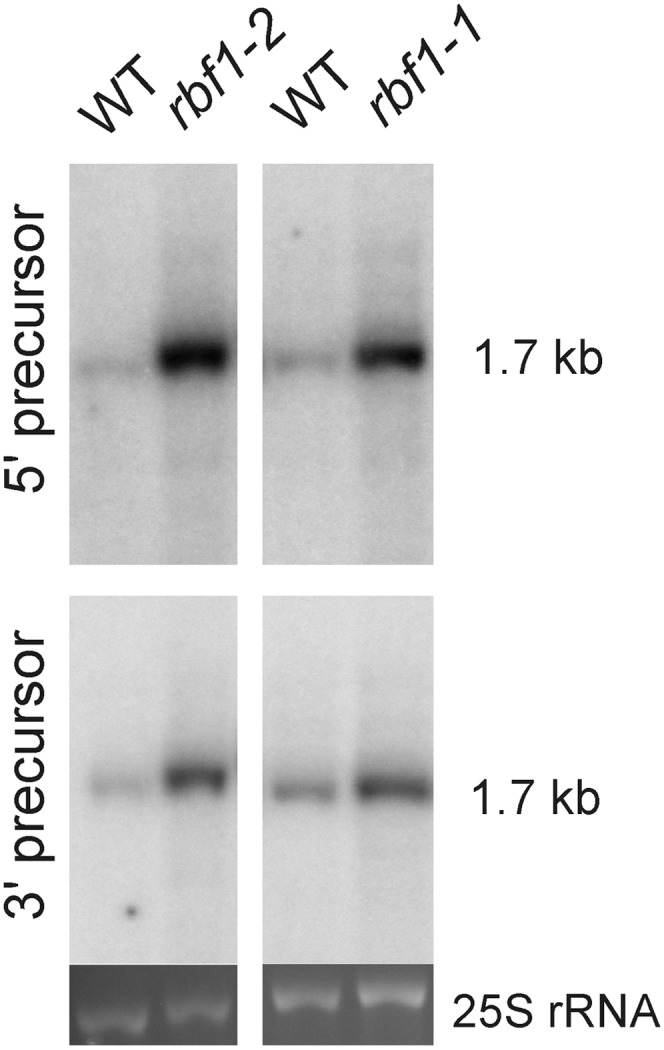
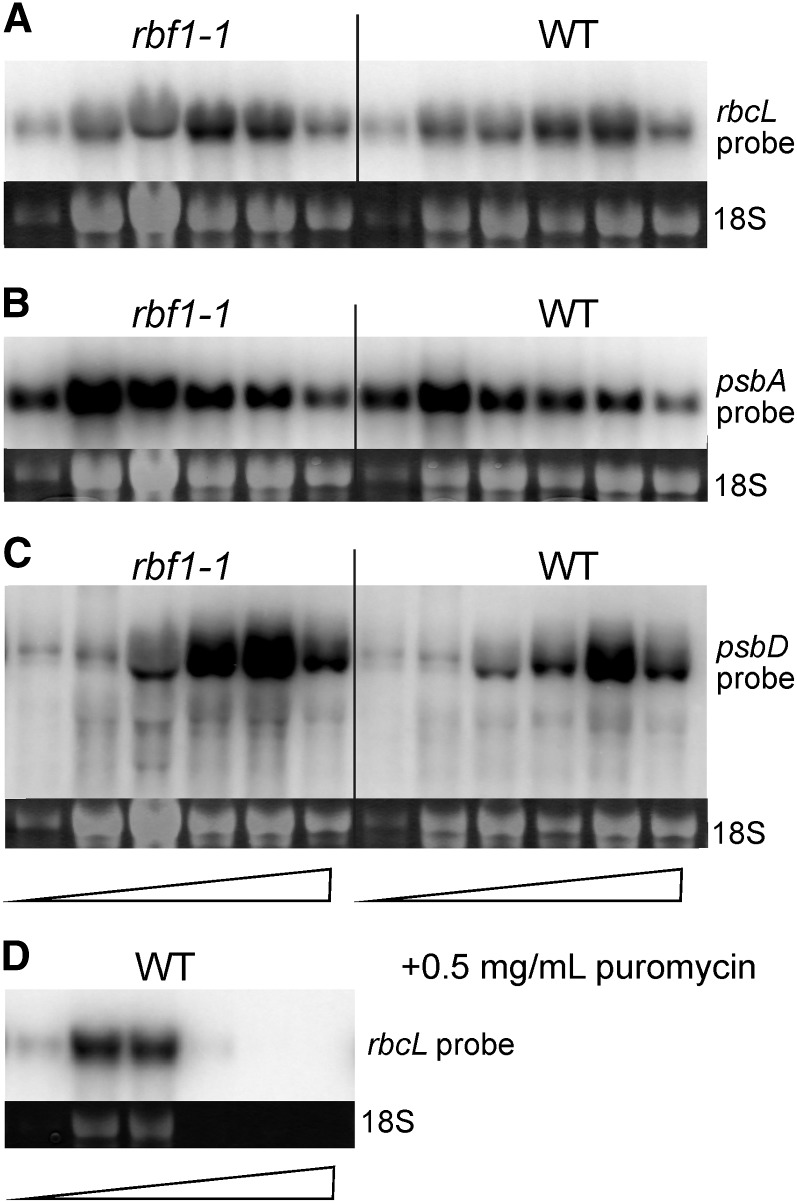
Plastids (chloroplasts) possess 70S ribosomes that are very similar in structure and function to the ribosomes of their bacterial ancestors. While most components of the bacterial ribosome (ribosomal RNAs [rRNAs] and ribosomal proteins) are well conserved in the plastid ribosome, little is known about the factors mediating the biogenesis of plastid ribosomes. Here, we have investigated a putative homolog of the bacterial RbfA (for ribosome-binding factor A) protein that was identified as a cold-shock protein and an auxiliary factor acting in the 5' maturation of the 16S rRNA. The unicellular green alga Chlamydomonas reinhardtii and the vascular plant Arabidopsis (Arabidopsis thaliana) both encode a single RbfA-like protein in their nuclear genomes. By generating specific antibodies against this protein, we show that the plant RbfA-like protein functions exclusively in the plastid, where it is associated with thylakoid membranes. Analysis of mutants for the corresponding gene (termed RBF1) reveals that the gene function is essential for photoautotrophic growth. Weak mutant alleles display reduced levels of plastid ribosomes, a specific depletion in 30S ribosomal subunits, and reduced activity of plastid protein biosynthesis. Our data suggest that, while the function in ribosome maturation and 16S rRNA 5' end processing is conserved, the RBF1 protein has assumed an additional role in 3' end processing. Together with the apparent absence of a homologous protein from plant mitochondria, our findings illustrate that the assembly process of the 70S ribosome is not strictly conserved and has undergone some modifications during organelle evolution.
Figures







References
-
- Albrecht V, Ingenfeld A, Apel K. (2006) Characterization of the snowy cotyledon 1 mutant of Arabidopsis thaliana: the impact of chloroplast elongation factor G on chloroplast development and plant vitality. Plant Mol Biol 60: 507–518 - PubMed
-
- Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408: 796–815 - PubMed
-
- Archibald JM. (2009) The puzzle of plastid evolution. Curr Biol 19: R81–R88 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
