

Human proT-cells generated in vitro facilitate hematopoietic stem cell-derived T-lymphopoiesis in vivo and restore thymic architecture
- PMID: 24215033
- PMCID: PMC5527400
- DOI: 10.1182/blood-2012-12-472803
Human proT-cells generated in vitro facilitate hematopoietic stem cell-derived T-lymphopoiesis in vivo and restore thymic architecture
Abstract
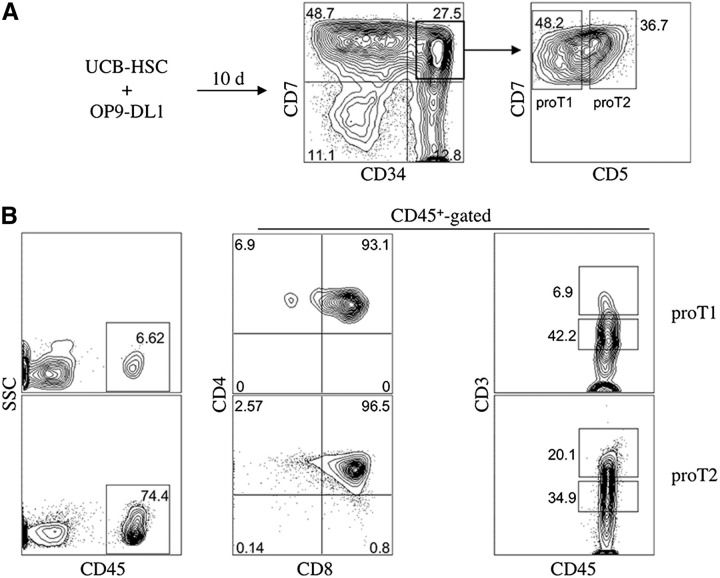
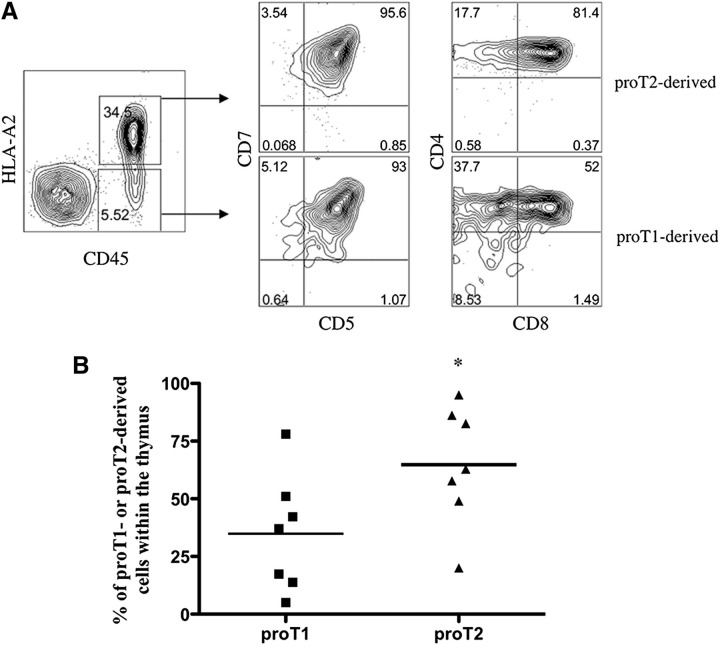
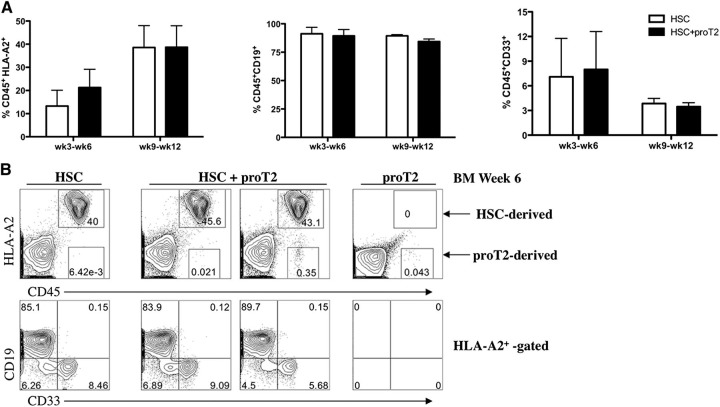
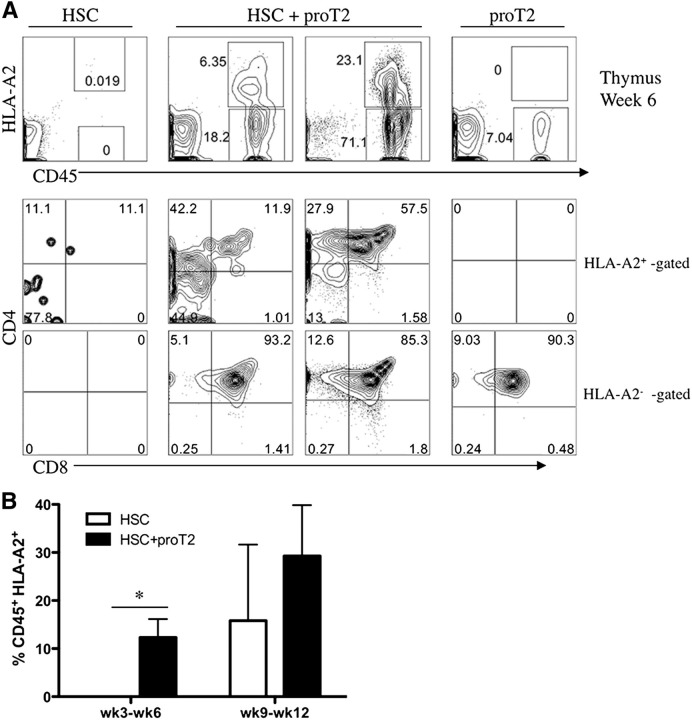
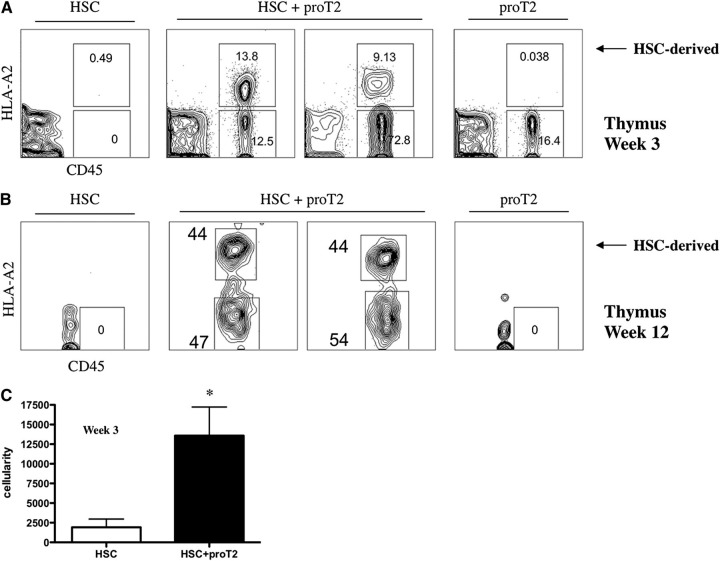
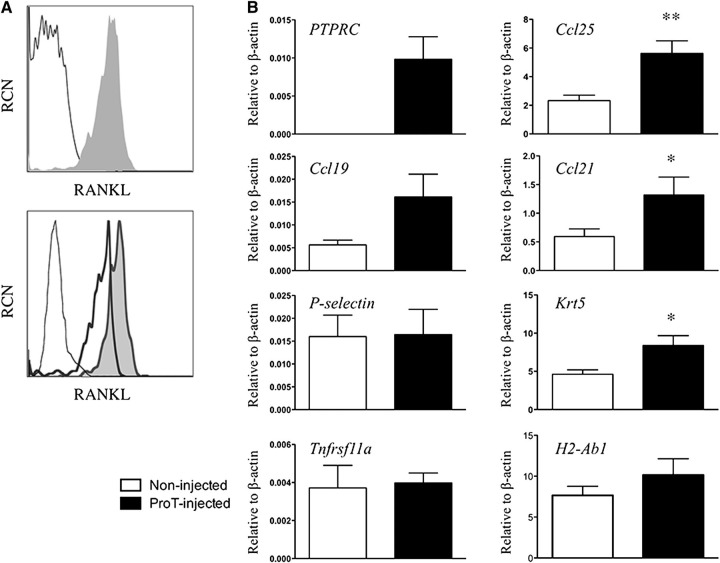
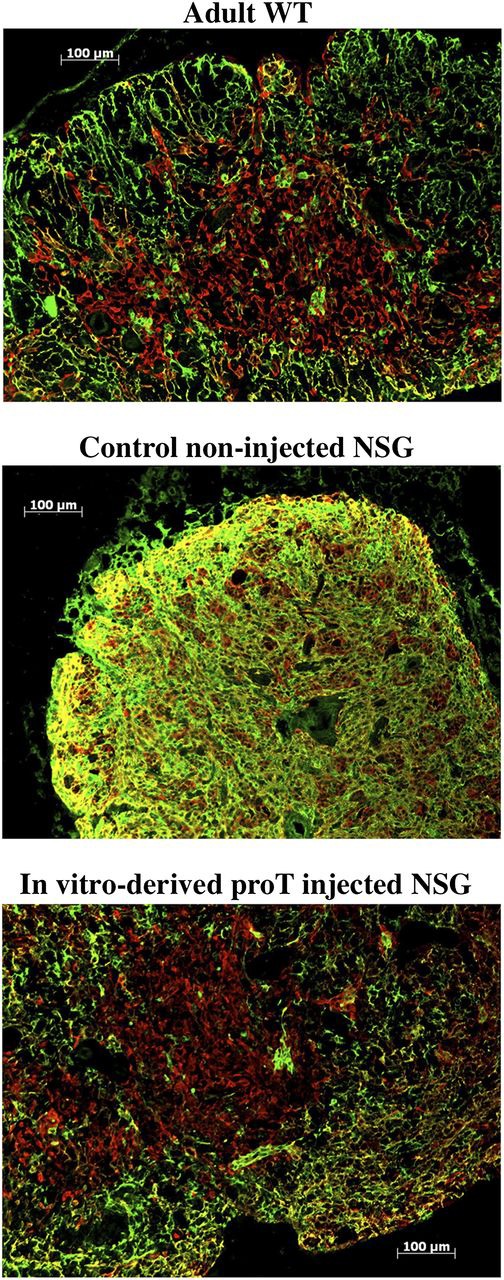
Hematopoietic stem cell transplantation (HSCT) is followed by a period of immune deficiency due to a paucity in T-cell reconstitution. Underlying causes are a severely dysfunctional thymus and an impaired production of thymus-seeding progenitors in the host. Here, we addressed whether in vitro-derived human progenitor T (proT)-cells could not only represent a source of thymus-seeding progenitors, but also able to influence the recovery of the thymic microenvironment. We examined whether co-transplantation of in vitro-derived human proT-cells with hematopoietic stem cells (HSCs) was able to facilitate HSC-derived T-lymphopoiesis posttransplant. A competitive transfer approach was used to define the optimal proT subset capable of reconstituting immunodeficient mice. Although the 2 subsets tested (proT1, CD34(+)CD7(+)CD5(-); proT2, CD34(+)CD7(+)CD5(+)) showed thymus engrafting function, proT2-cells exhibited superior engrafting capacity. Based on this, when proT2-cells were coinjected with HSCs, a significantly improved and accelerated HSC-derived T-lymphopoiesis was observed. Furthermore, we uncovered a potential mechanism by which receptor activator of nuclear factor κb (RANK) ligand-expressing proT2-cells induce changes in both the function and architecture of the thymus microenvironment, which favors the recruitment of bone marrow-derived lymphoid progenitors. Our findings provide further support for the use of Notch-expanded progenitors in cell-based therapies to aid in the recovery of T-cells in patients undergoing HSCT.
Figures
References
-
- Holländer G, Gill J, Zuklys S, Iwanami N, Liu C, Takahama Y. Cellular and molecular events during early thymus development. Immunol Rev. 2006;209:28–46. - PubMed
-
- Blackburn CC, Manley NR. Developing a new paradigm for thymus organogenesis. Nat Rev Immunol. 2004;4(4):278–289. - PubMed
-
- Petrie HT, Zúñiga-Pflücker JC. Zoned out: functional mapping of stromal signaling microenvironments in the thymus. Annu Rev Immunol. 2007;25:649–679. - PubMed
-
- Shortman K, Wu L. Early T lymphocyte progenitors. Annu Rev Immunol. 1996;14:29–47. - PubMed
-
- Goldschneider I. Cyclical mobilization and gated importation of thymocyte progenitors in the adult mouse: evidence for a thymus-bone marrow feedback loop. Immunol Rev. 2006;209:58–75. - PubMed
Publication types
MeSH terms
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
