

Human-specific endogenous retroviral insert serves as an enhancer for the schizophrenia-linked gene PRODH
- PMID: 24218577
- PMCID: PMC3845128
- DOI: 10.1073/pnas.1318172110
Human-specific endogenous retroviral insert serves as an enhancer for the schizophrenia-linked gene PRODH
Abstract
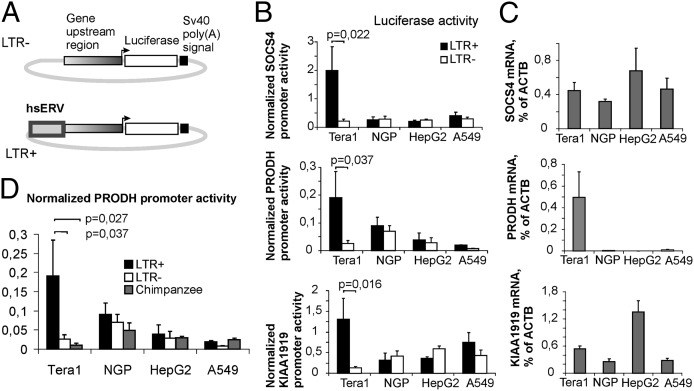
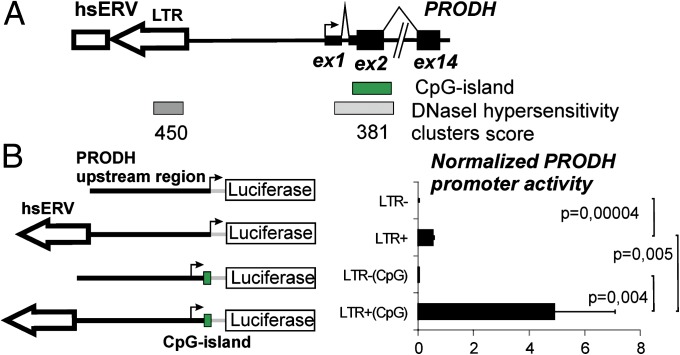
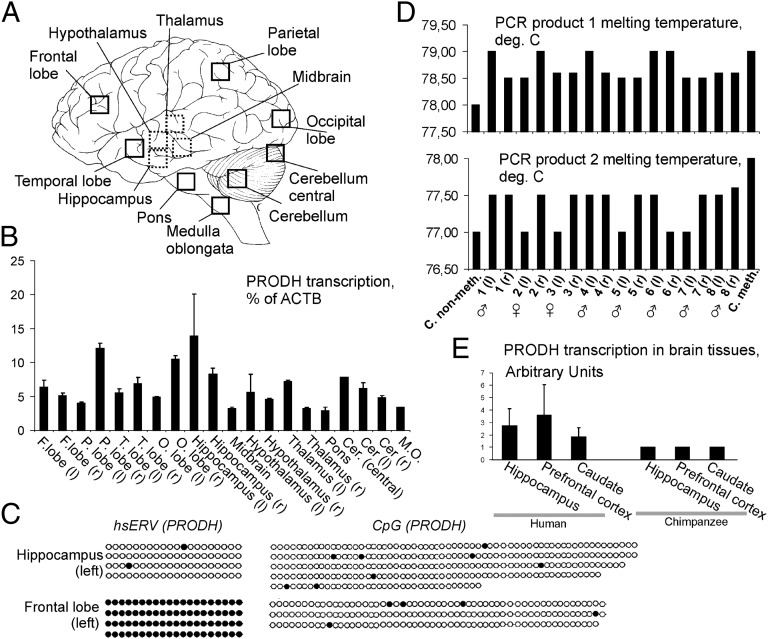
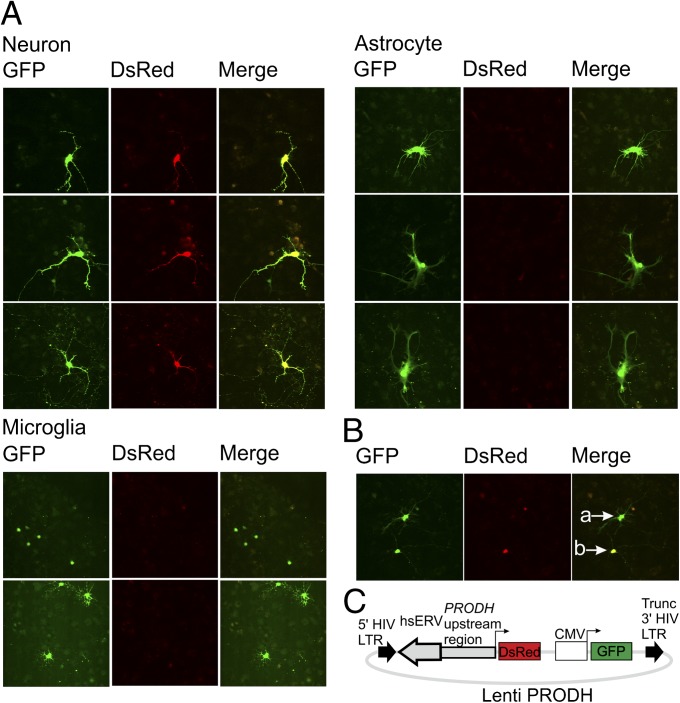
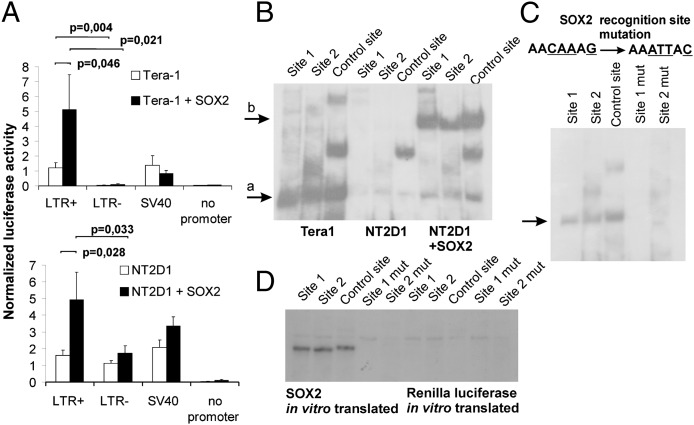
Using a systematic, whole-genome analysis of enhancer activity of human-specific endogenous retroviral inserts (hsERVs), we identified an element, hsERVPRODH, that acts as a tissue-specific enhancer for the PRODH gene, which is required for proper CNS functioning. PRODH is one of the candidate genes for susceptibility to schizophrenia and other neurological disorders. It codes for a proline dehydrogenase enzyme, which catalyses the first step of proline catabolism and most likely is involved in neuromediator synthesis in the CNS. We investigated the mechanisms that regulate hsERVPRODH enhancer activity. We showed that the hsERVPRODH enhancer and the internal CpG island of PRODH synergistically activate its promoter. The enhancer activity of hsERVPRODH is regulated by methylation, and in an undermethylated state it can up-regulate PRODH expression in the hippocampus. The mechanism of hsERVPRODH enhancer activity involves the binding of the transcription factor SOX2, whch is preferentially expressed in hippocampus. We propose that the interaction of hsERVPRODH and PRODH may have contributed to human CNS evolution.
Keywords: DNA methylation; central nervous system; human speciation; human-specific endogenous retrovirus; retroelement.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
