

Differential effects of global versus local testosterone on singing behavior and its underlying neural substrate
- PMID: 24218603
- PMCID: PMC3845175
- DOI: 10.1073/pnas.1311371110
Differential effects of global versus local testosterone on singing behavior and its underlying neural substrate
Abstract
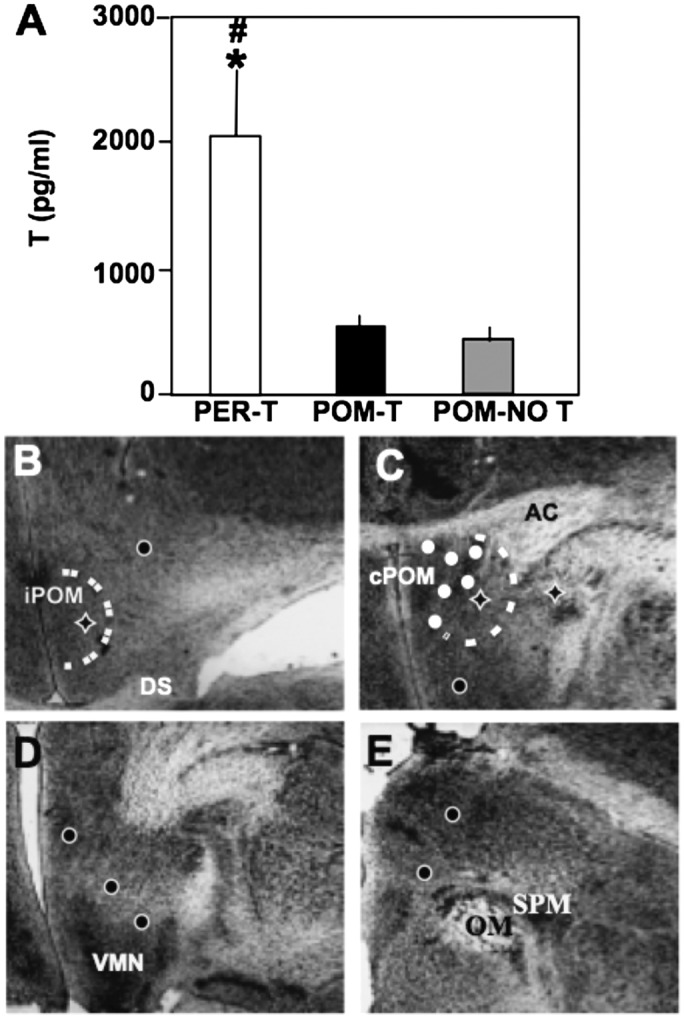
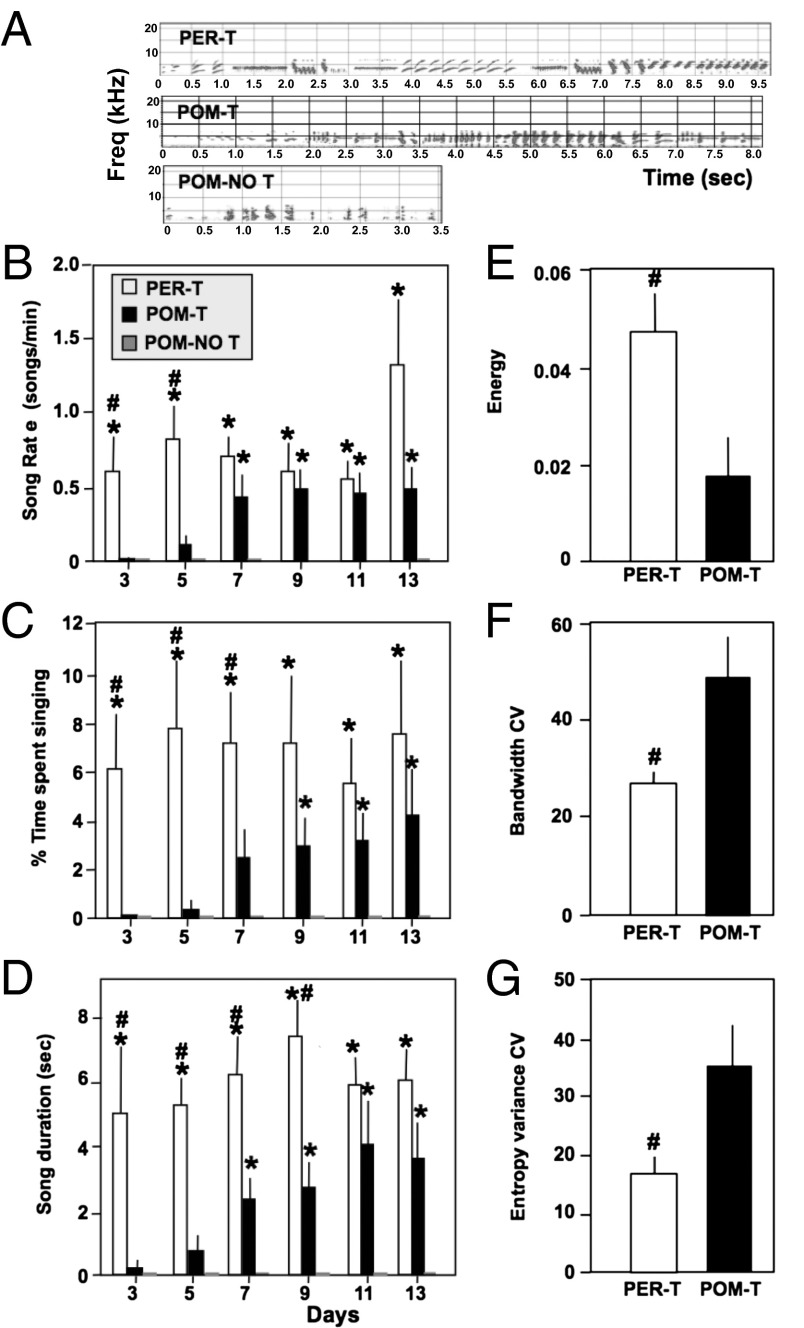
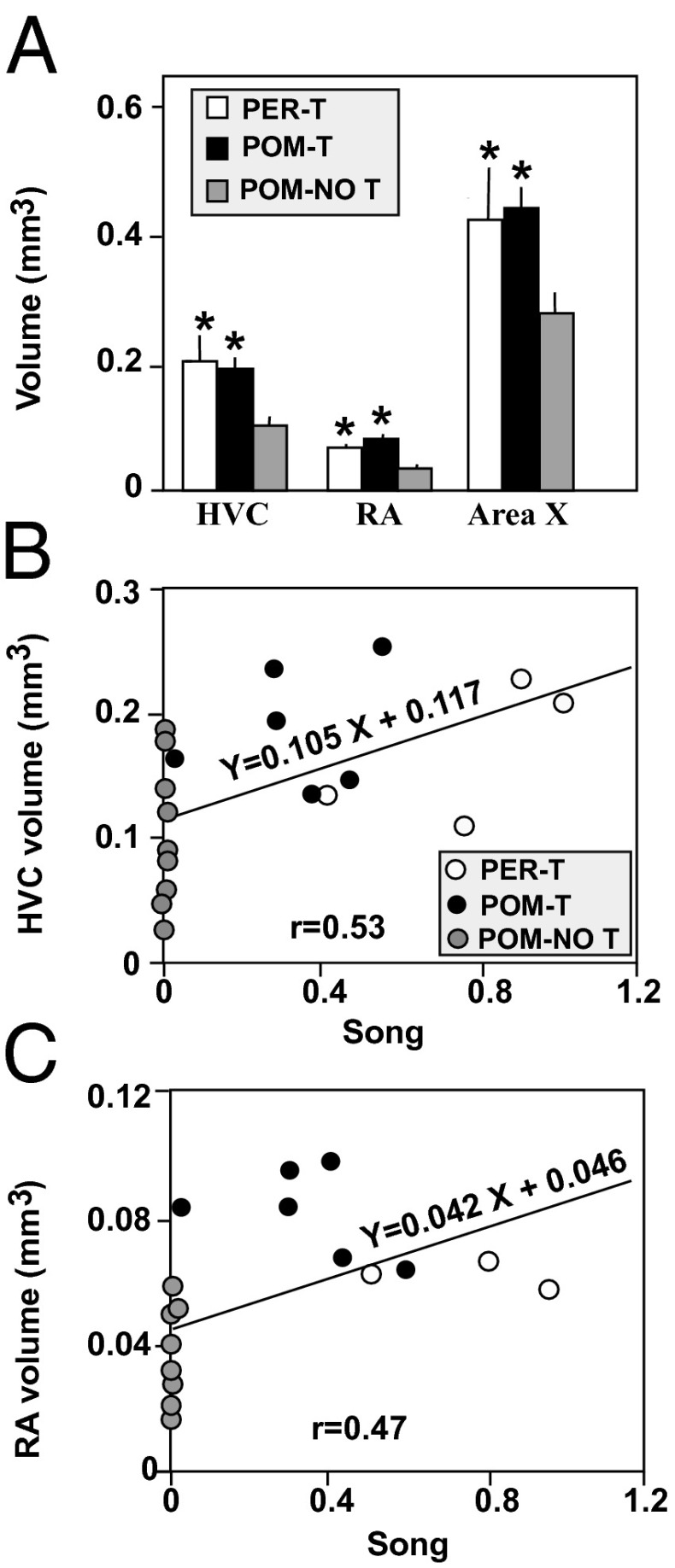
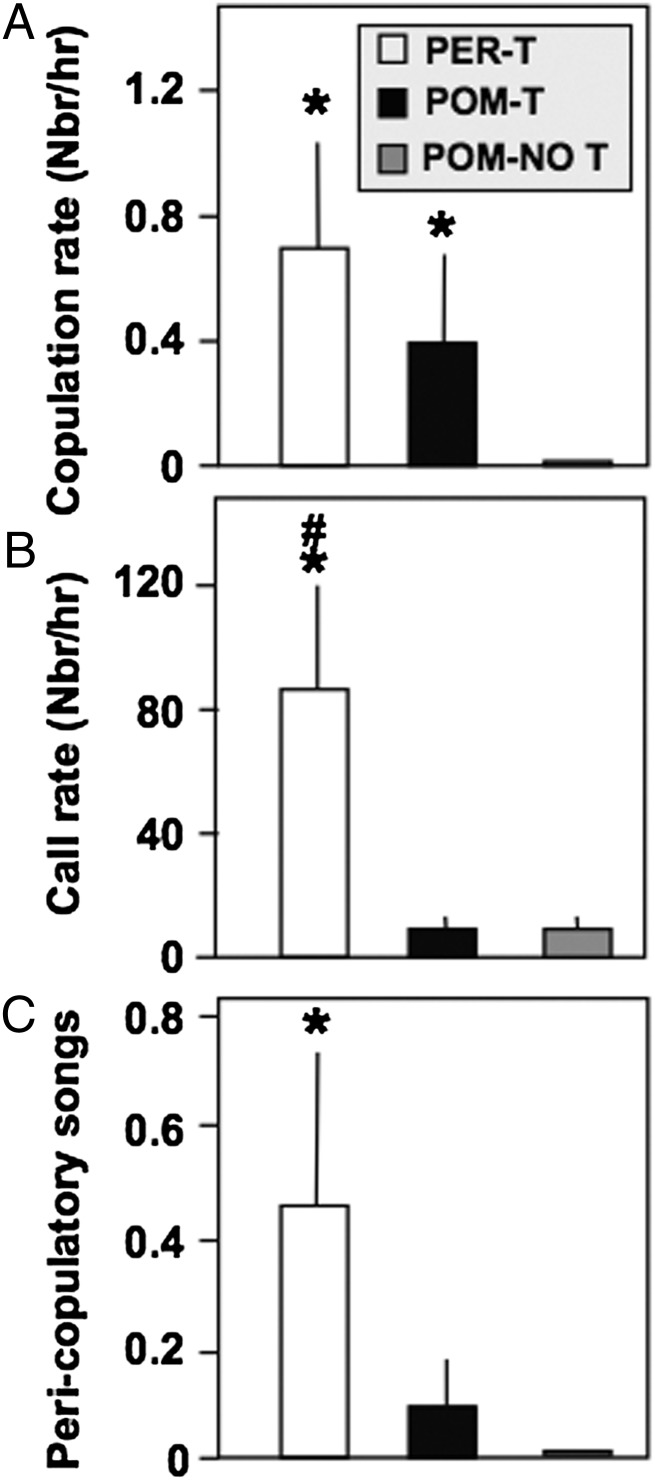
Steroid hormones regulate multiple but distinct aspects of social behaviors. Testosterone (T) has multiple effects on learned courtship song in that it regulates both the motivation to sing in a particular social context as well as the quality of song produced. The neural substrate(s) where T acts to regulate the motivation to sing as opposed to other aspects of song has not been definitively characterized. We show here that T implants in the medial preoptic nucleus (POM) of castrated male canaries (Serinus canaria) increase song rate but do not enhance acoustic features such as song stereotypy compared with birds receiving peripheral T that can act globally throughout the brain. Strikingly, T action in the POM increased song control nuclei volume, consistent with the hypothesis that singing activity induces neuroplasticity in the song control system independent of T acting in these nuclei. When presented with a female canary, POM-T birds copulated at a rate comparable to birds receiving systemic T but produced fewer calls and songs in her presence. Thus, POM is a key site where T acts to activate copulation and increase song rate, an appetitive sexual behavior in songbirds, but T action in other areas of the brain or periphery (e.g., HVC, dopaminergic cell groups, or the syrinx) is required to enhance the quality of song (i.e., stereotypy) as well as regulate context-specific vocalizations. These results have broad implications for research concerning how steroids act at multiple brain loci to regulate distinct sociosexual behaviors and the associated neuroplasticity.
Keywords: activity-driven plasticity; preoptic area; singing motivation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Adkins-Regan E. Neuroendocrinology of social behavior. ILAR J. 2009;50(1):5–14. - PubMed
-
- Ball GF, Riters LV, Balthazart J. Neuroendocrinology of song behavior and avian brain plasticity: Multiple sites of action of sex steroid hormones. Front Neuroendocrinol. 2002;23(2):137–178. - PubMed
-
- Bottjer SW, Johnson F. Circuits, hormones, and learning: Vocal behavior in songbirds. J Neurobiol. 1997;33(5):602–618. - PubMed
-
- Lee AW, Pfaff DW. Hormone effects on specific and global brain functions. J Physiol Sci. 2008;58(4):213–220. - PubMed
-
- Arnold AP. Logical levels of steroid hormone action in the control of vertebrate behavior. American Zoologist. 1981;21:233–242.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
