

Structural and functional insights into the regulation mechanism of CK2 by IP6 and the intrinsically disordered protein Nopp140
- PMID: 24218616
- PMCID: PMC3845154
- DOI: 10.1073/pnas.1304670110
Structural and functional insights into the regulation mechanism of CK2 by IP6 and the intrinsically disordered protein Nopp140
Abstract
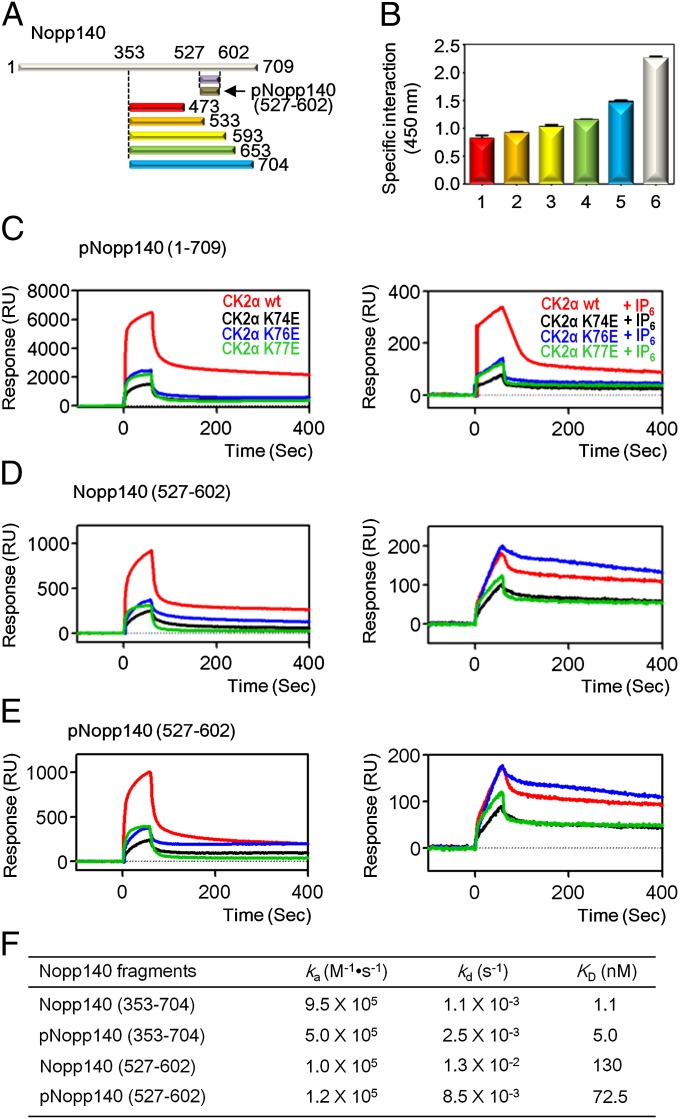
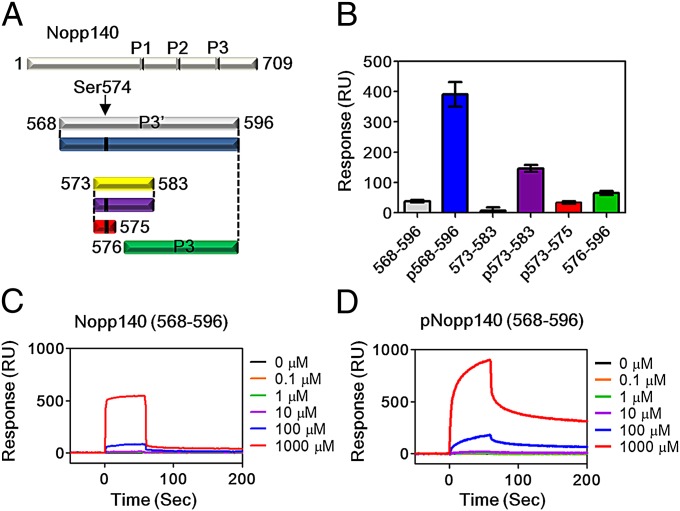
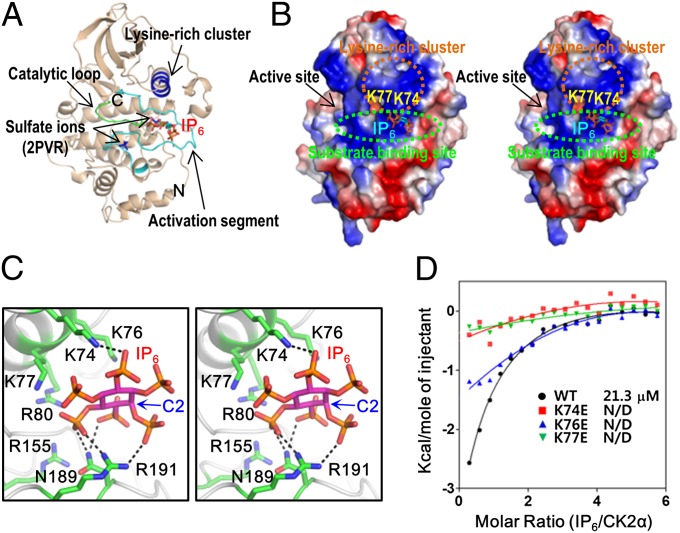
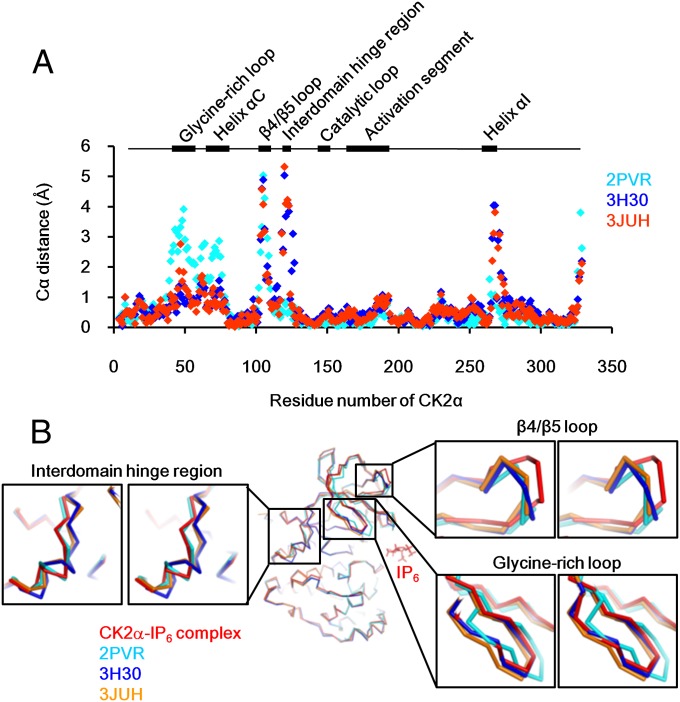
Protein kinase CK2 is a ubiquitous kinase that can phosphorylate hundreds of cellular proteins and plays important roles in cell growth and development. Deregulation of CK2 is related to a variety of human cancers, and CK2 is regarded as a suppressor of apoptosis; therefore, it is a target of anticancer therapy. Nucleolar phosphoprotein 140 (Nopp140), which is an intrinsically disordered protein, interacts with CK2 and inhibits the latter's catalytic activity in vitro. Interestingly, the catalytic activity of CK2 is recovered in the presence of d-myo-inositol 1,2,3,4,5,6-hexakisphosphate (IP6). IP6 is widely distributed in animal cells, but the molecular mechanisms that govern its cellular functions in animal cells have not been completely elucidated. In this study, the crystal structure of CK2 in complex with IP6 showed that the lysine-rich cluster of CK2 plays an important role in binding to IP6. The biochemical experiments revealed that a Nopp140 fragment (residues 568-596) and IP6 competitively bind to the catalytic subunit of CK2 (CK2α), and phospho-Ser574 of Nopp140 significantly enhances its interaction with CK2α. Substitutions of K74E, K76E, and K77E in CK2α significantly reduced the interactions of CK2α with both IP6 and the Nopp140-derived peptide. Our study gives an insight into the regulation of CK2. In particular, our work suggests that CK2 activity is inhibited by Nopp140 and reactivated by IP6 by competitive binding at the substrate recognition site of CK2.
Keywords: IDP; inositol hexakisphosphate; phosphorylation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
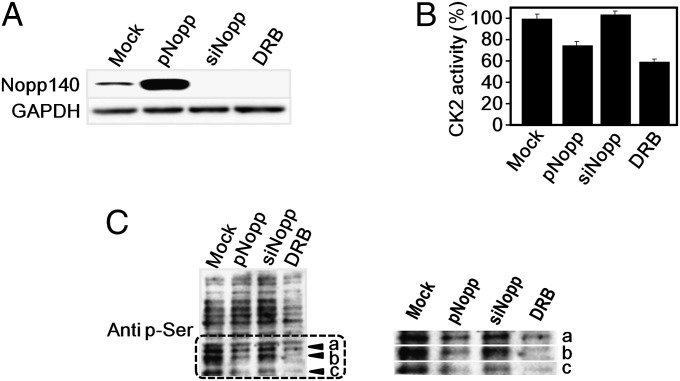
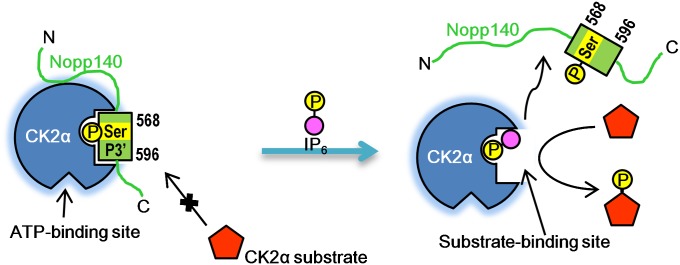
References
-
- Tawfic S, et al. Protein kinase CK2 signal in neoplasia. Histol Histopathol. 2001;16(2):573–582. - PubMed
-
- Seldin DC, Leder P. Casein kinase II alpha transgene-induced murine lymphoma: Relation to theileriosis in cattle. Science. 1995;267(5199):894–897. - PubMed
-
- Ahmed K, et al. Significance of protein kinase CK2 nuclear signaling in neoplasia. J Cell Biochem Suppl. 2000;35(Suppl 35):130–135. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
