

Low-affinity transcription factor binding sites shape morphogen responses and enhancer evolution
- PMID: 24218631
- PMCID: PMC3826492
- DOI: 10.1098/rstb.2013.0018
Low-affinity transcription factor binding sites shape morphogen responses and enhancer evolution
Abstract
In the era of functional genomics, the role of transcription factor (TF)-DNA binding affinity is of increasing interest: for example, it has recently been proposed that low-affinity genomic binding events, though frequent, are functionally irrelevant. Here, we investigate the role of binding site affinity in the transcriptional interpretation of Hedgehog (Hh) morphogen gradients. We noted that enhancers of several Hh-responsive Drosophila genes have low predicted affinity for Ci, the Gli family TF that transduces Hh signalling in the fly. Contrary to our initial hypothesis, improving the affinity of Ci/Gli sites in enhancers of dpp, wingless and stripe, by transplanting optimal sites from the patched gene, did not result in ectopic responses to Hh signalling. Instead, we found that these enhancers require low-affinity binding sites for normal activation in regions of relatively low signalling. When Ci/Gli sites in these enhancers were altered to improve their binding affinity, we observed patterning defects in the transcriptional response that are consistent with a switch from Ci-mediated activation to Ci-mediated repression. Synthetic transgenic reporters containing isolated Ci/Gli sites confirmed this finding in imaginal discs. We propose that the requirement for gene activation by Ci in the regions of low-to-moderate Hh signalling results in evolutionary pressure favouring weak binding sites in enhancers of certain Hh target genes.
Keywords: Ci/Gli; binding affinity; enhancers; gene regulation; hedgehog signalling; morphogen gradient.
Figures
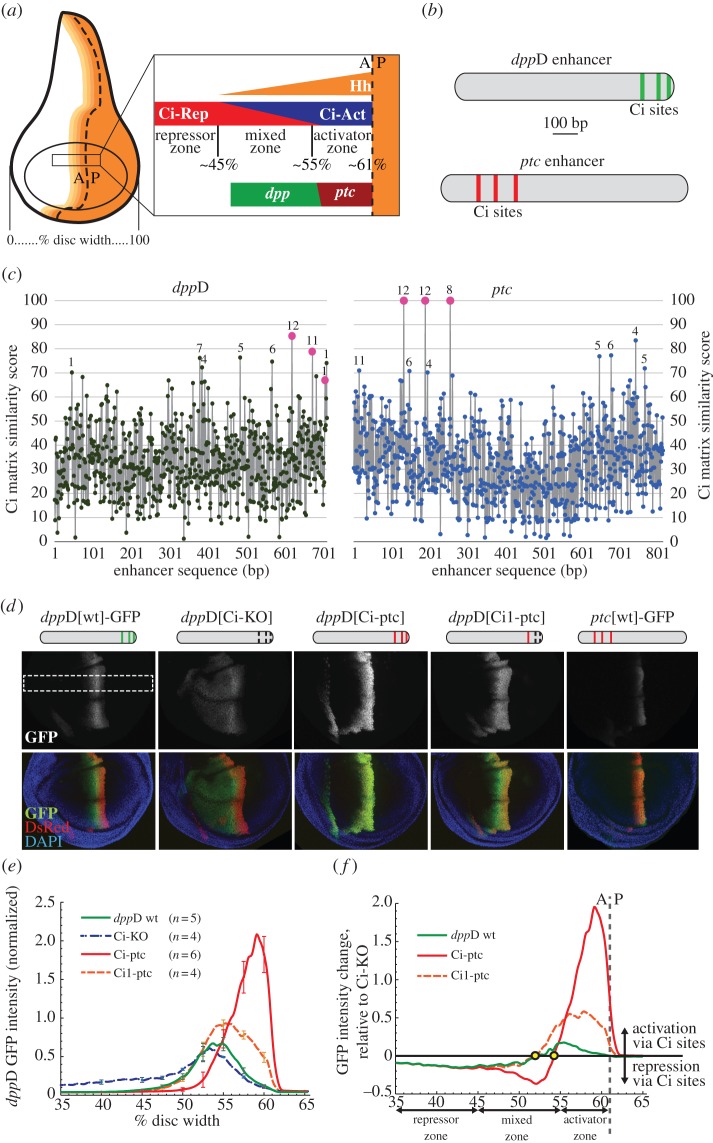
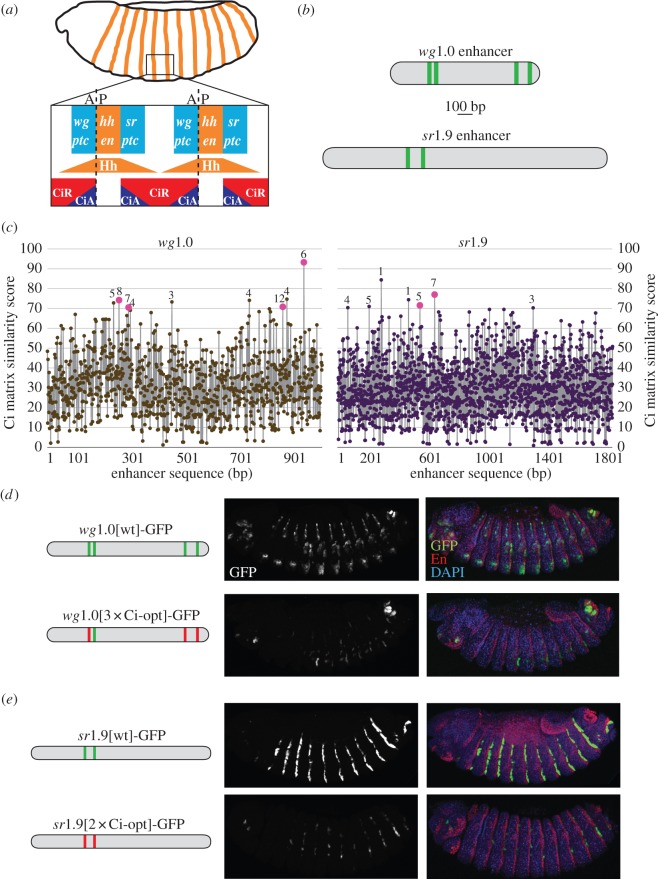

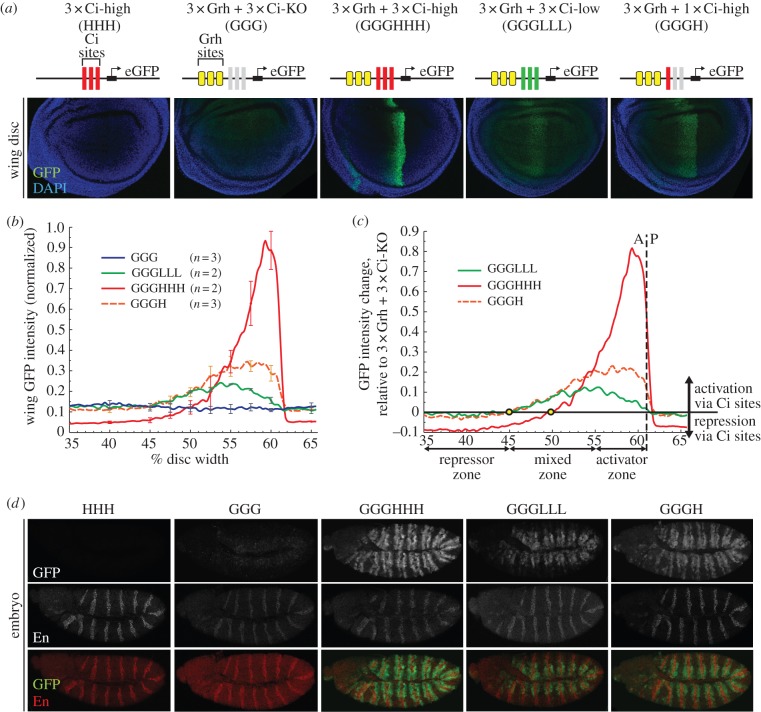
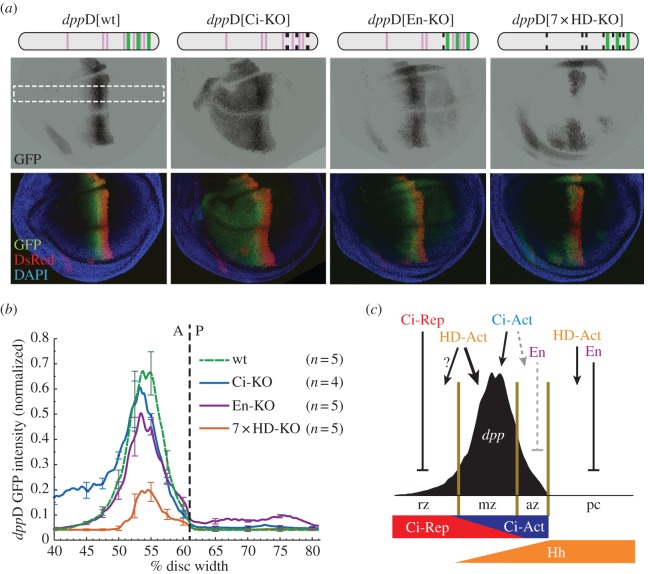
References
-
- Levine M. 2010. Transcriptional enhancers in animal development and evolution. Curr. Biol. 20, R754–R763 (doi:10.1016/j.cub.2010.06.070) - DOI - PMC - PubMed
-
- Stamatoyannopoulos JA. 2012. What does our genome encode? Genome Res. 22, 1602–1611 (doi:10.1101/gr.146506.112) - DOI - PMC - PubMed
-
- Ong C-T, Corces VG. 2011. Enhancer function: new insights into the regulation of tissue-specific gene expression. Nat. Rev. Genet. 12, 283–293 (doi:10.1038/nrg2957) - DOI - PMC - PubMed
-
- Göke J, Jung M, Behrens S, Chavez L, O'Keeffe S, Timmermann B, Lehrach H, Adjaye J, Vingron M. 2011. Combinatorial binding in human and mouse embryonic stem cells identifies conserved enhancers active in early embryonic development. PLoS Comput. Biol. 7, e1002304 (doi:10.1371/journal.pcbi.1002304) - DOI - PMC - PubMed
-
- Shen Y, et al. 2012. A map of the cis-regulatory sequences in the mouse genome. Nature 488, 116–120 (doi:10.1038/nature11243) - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
