

Identification of a pan-cancer oncogenic microRNA superfamily anchored by a central core seed motif
- PMID: 24220575
- PMCID: PMC3868236
- DOI: 10.1038/ncomms3730
Identification of a pan-cancer oncogenic microRNA superfamily anchored by a central core seed motif
Abstract
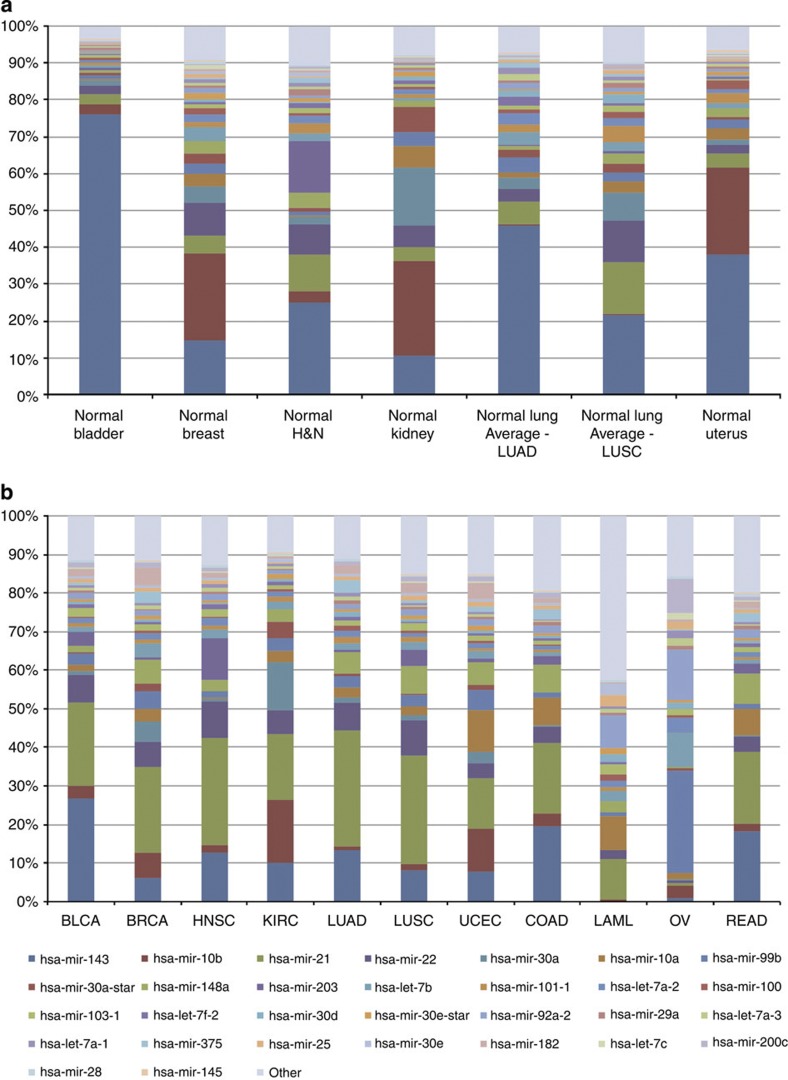
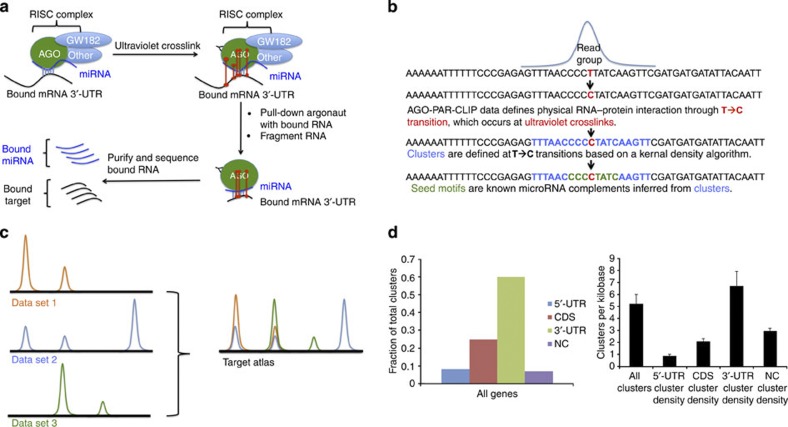
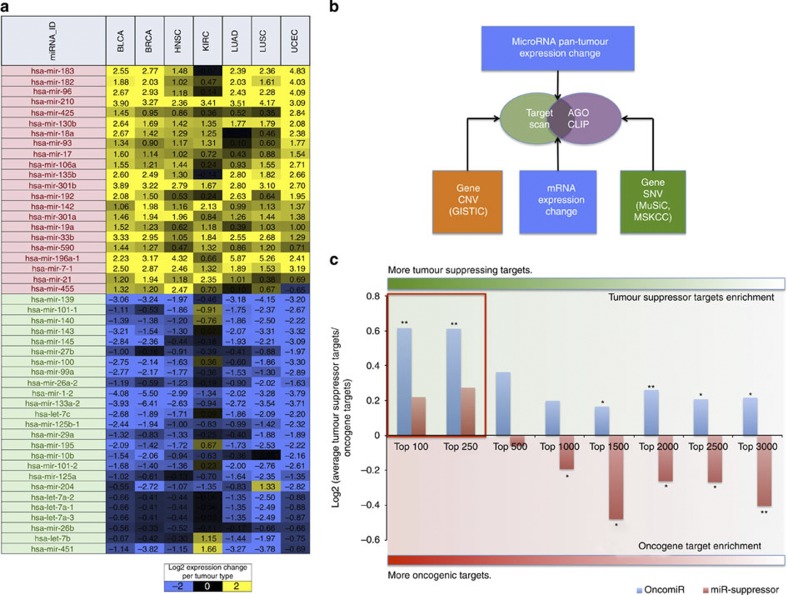
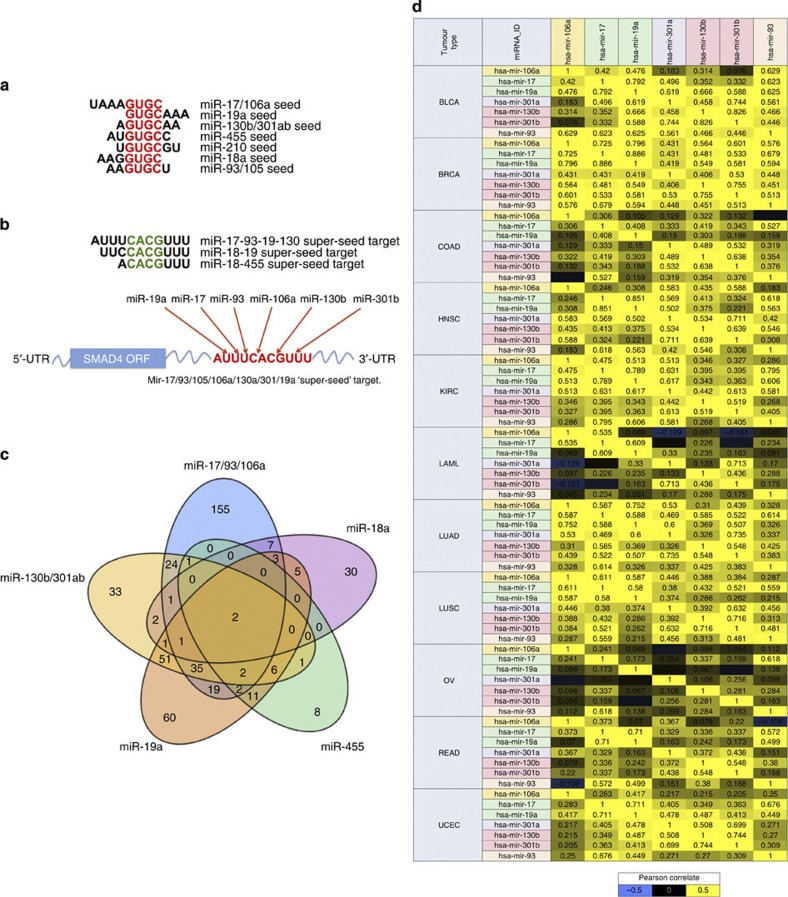
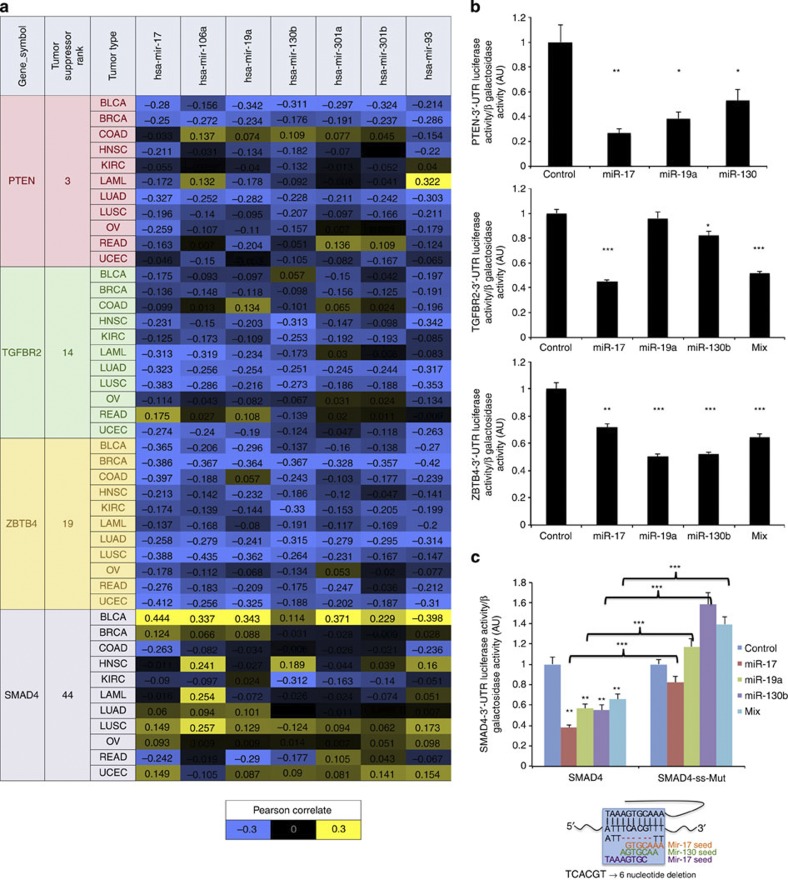
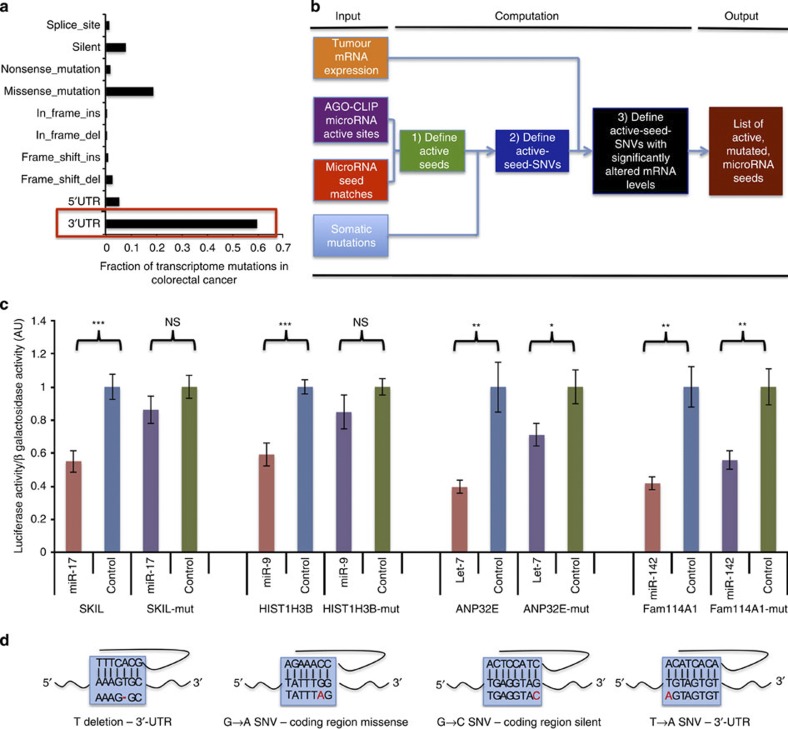
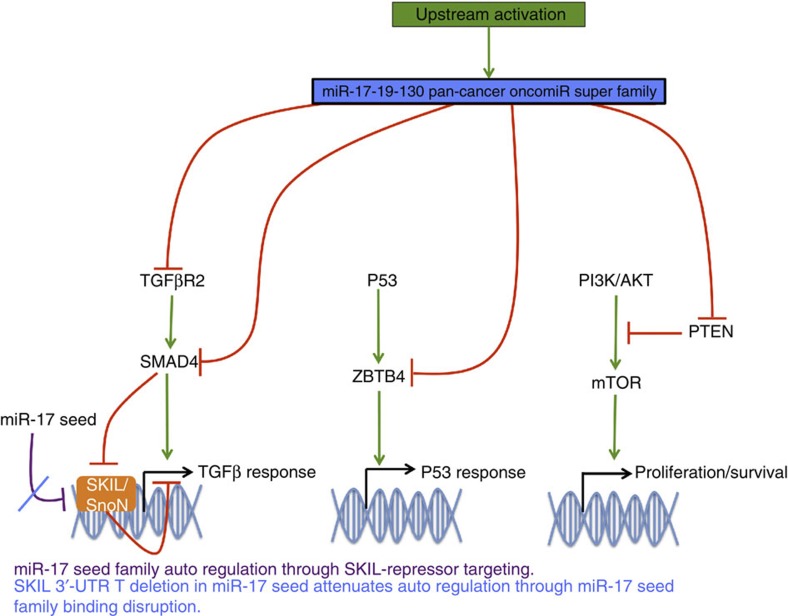
MicroRNAs modulate tumorigenesis through suppression of specific genes. As many tumour types rely on overlapping oncogenic pathways, a core set of microRNAs may exist, which consistently drives or suppresses tumorigenesis in many cancer types. Here we integrate The Cancer Genome Atlas (TCGA) pan-cancer data set with a microRNA target atlas composed of publicly available Argonaute Crosslinking Immunoprecipitation (AGO-CLIP) data to identify pan-tumour microRNA drivers of cancer. Through this analysis, we show a pan-cancer, coregulated oncogenic microRNA 'superfamily' consisting of the miR-17, miR-19, miR-130, miR-93, miR-18, miR-455 and miR-210 seed families, which cotargets critical tumour suppressors via a central GUGC core motif. We subsequently define mutations in microRNA target sites using the AGO-CLIP microRNA target atlas and TCGA exome-sequencing data. These combined analyses identify pan-cancer oncogenic cotargeting of the phosphoinositide 3-kinase, TGFβ and p53 pathways by the miR-17-19-130 superfamily members.
Figures
References
-
- Selbach M. et al.. Widespread changes in protein synthesis induced by microRNAs. Nature 455, 58–63 (2008). - PubMed
-
- Lu J. et al.. MicroRNA expression profiles classify human cancers. Nature 435, 834–838 (2005). - PubMed
-
- Martello G. et al.. A microRNA targeting dicer for metastasis control. Cell 141, 1195–1207 (2010). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
