

Wnt7b is an important intrinsic regulator of hair follicle stem cell homeostasis and hair follicle cycling
- PMID: 24222445
- PMCID: PMC4398394
- DOI: 10.1002/stem.1599
Wnt7b is an important intrinsic regulator of hair follicle stem cell homeostasis and hair follicle cycling
Abstract
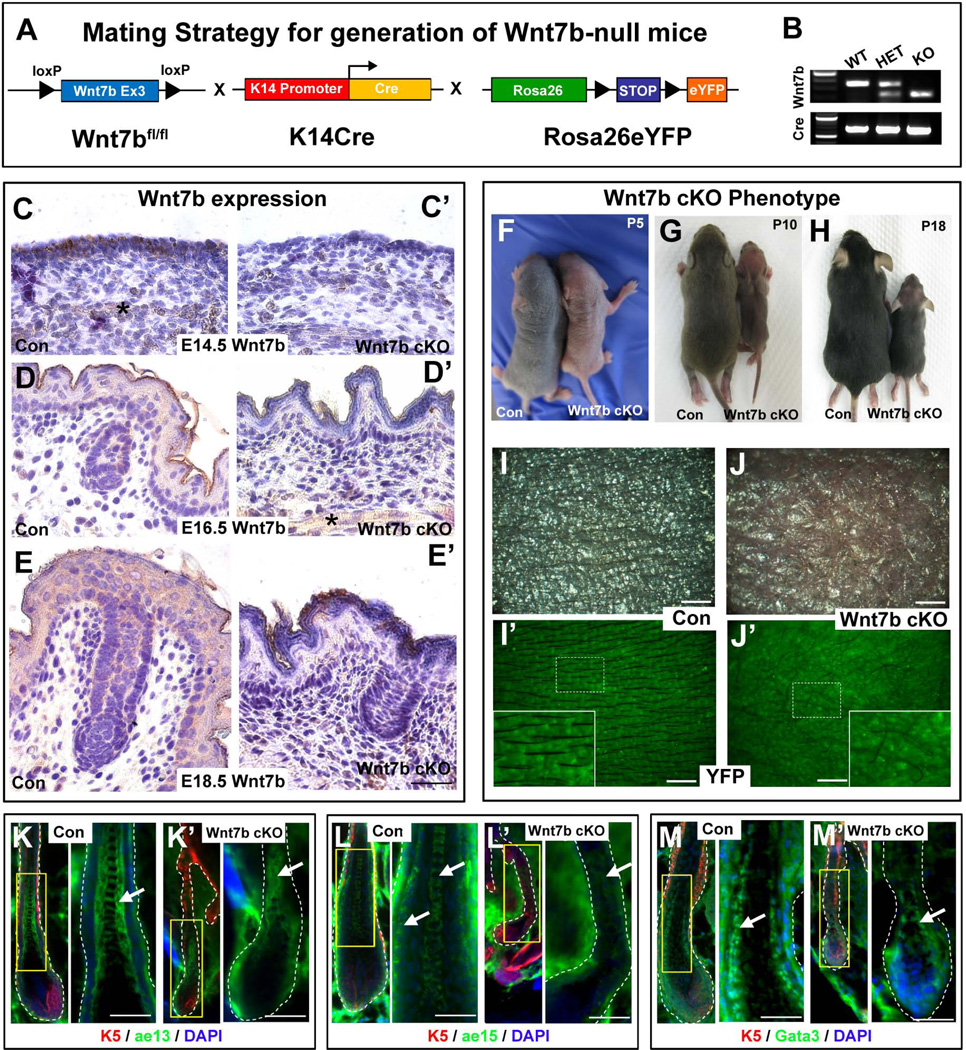
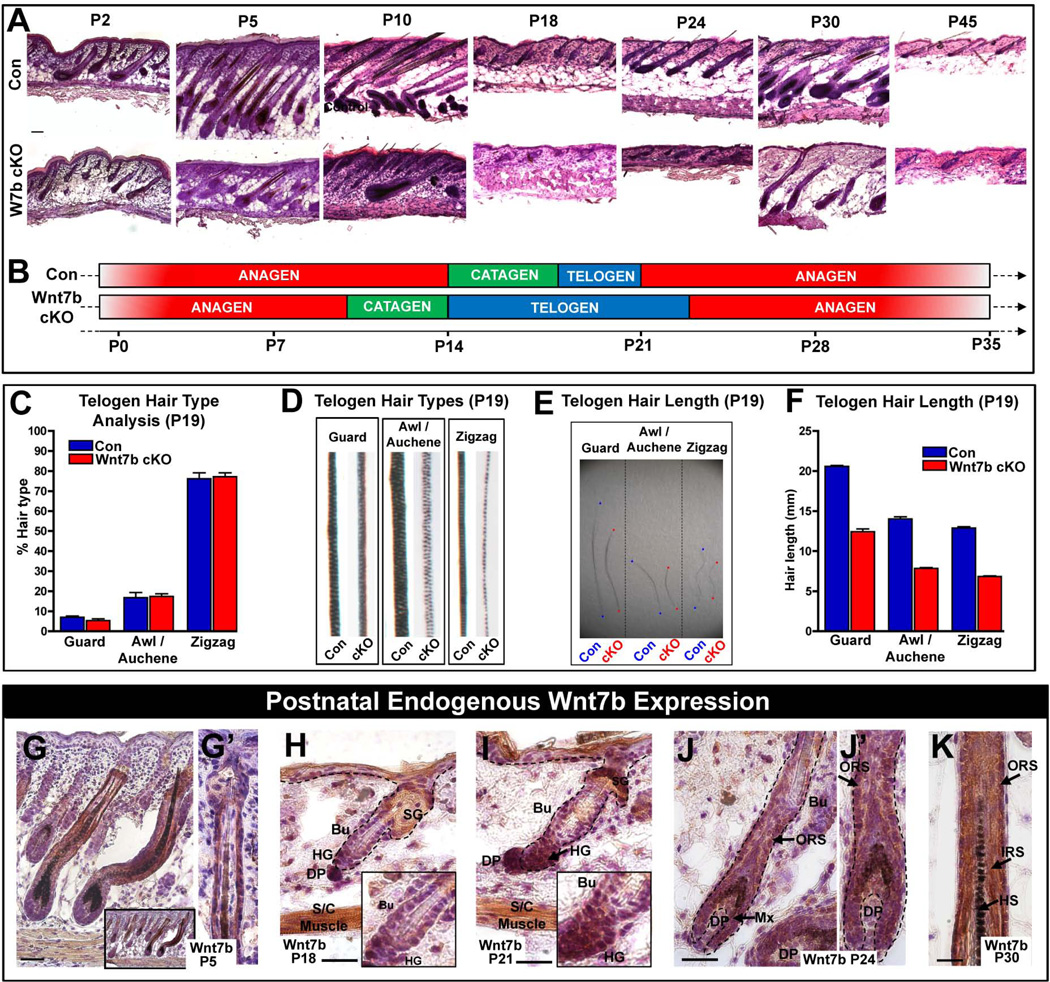
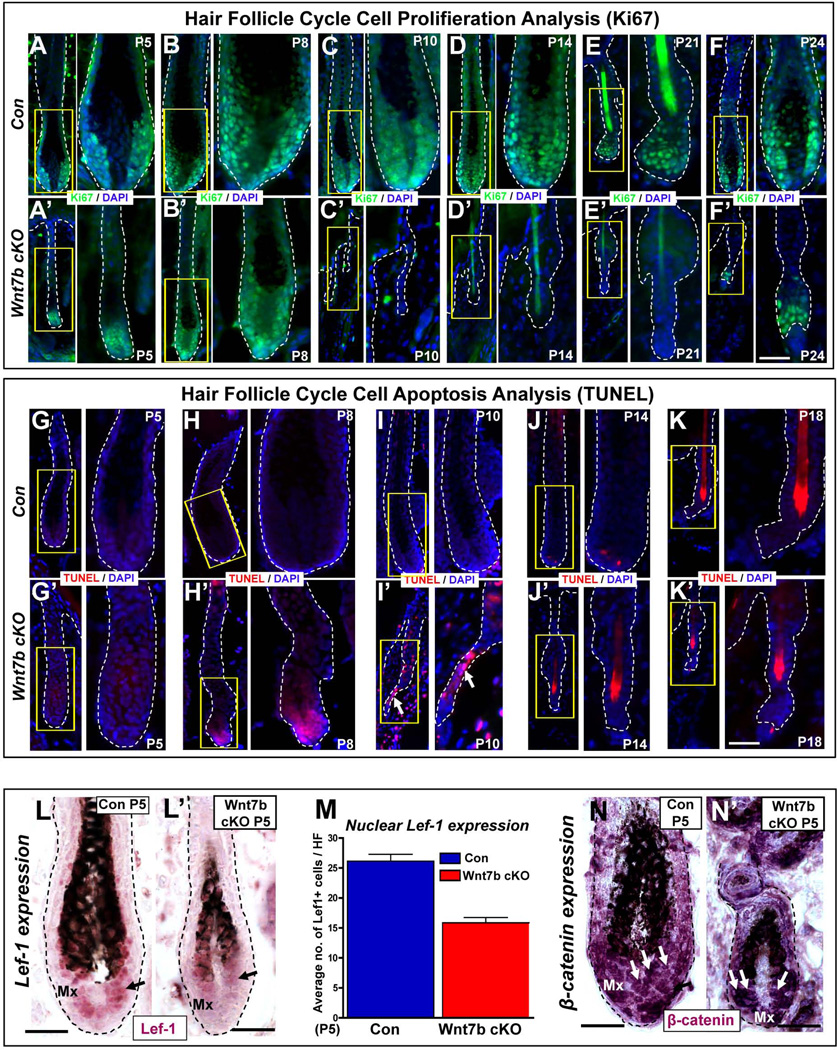
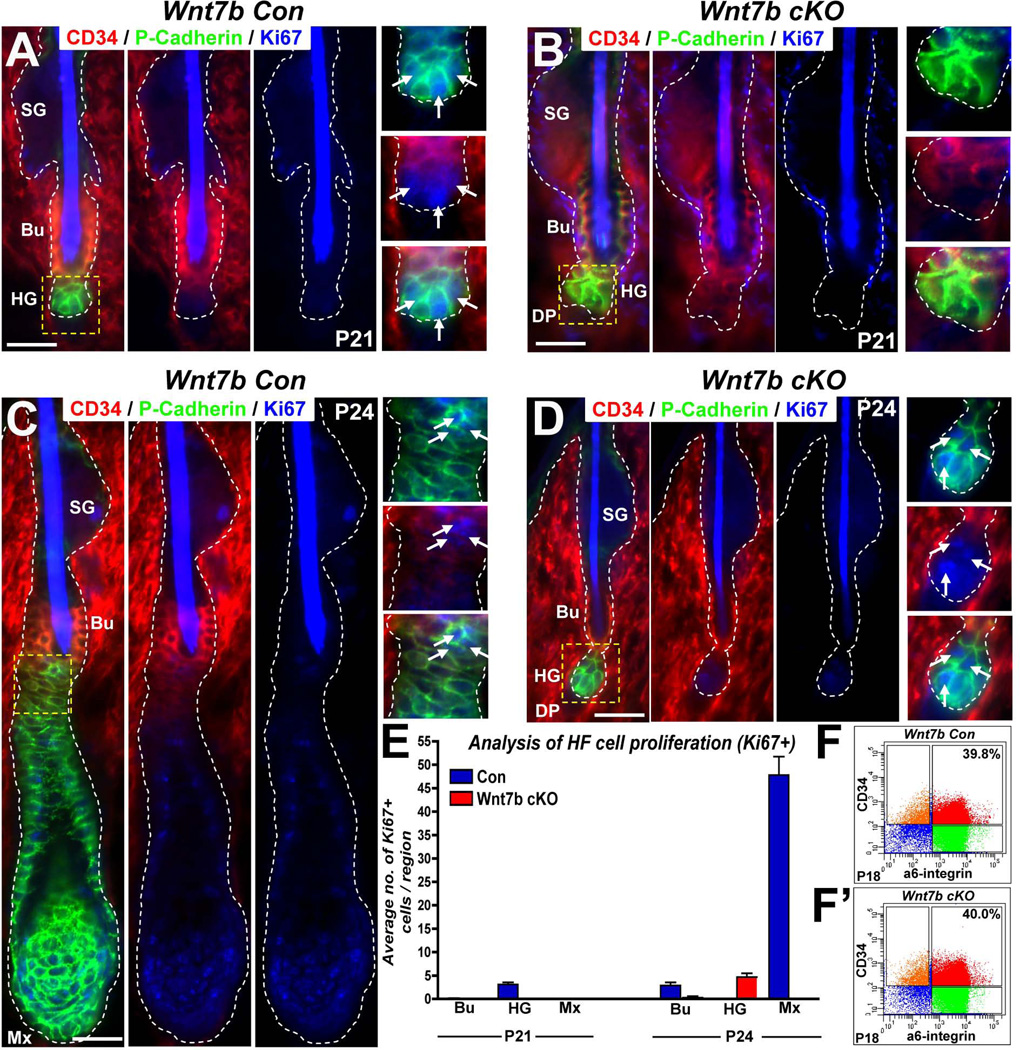
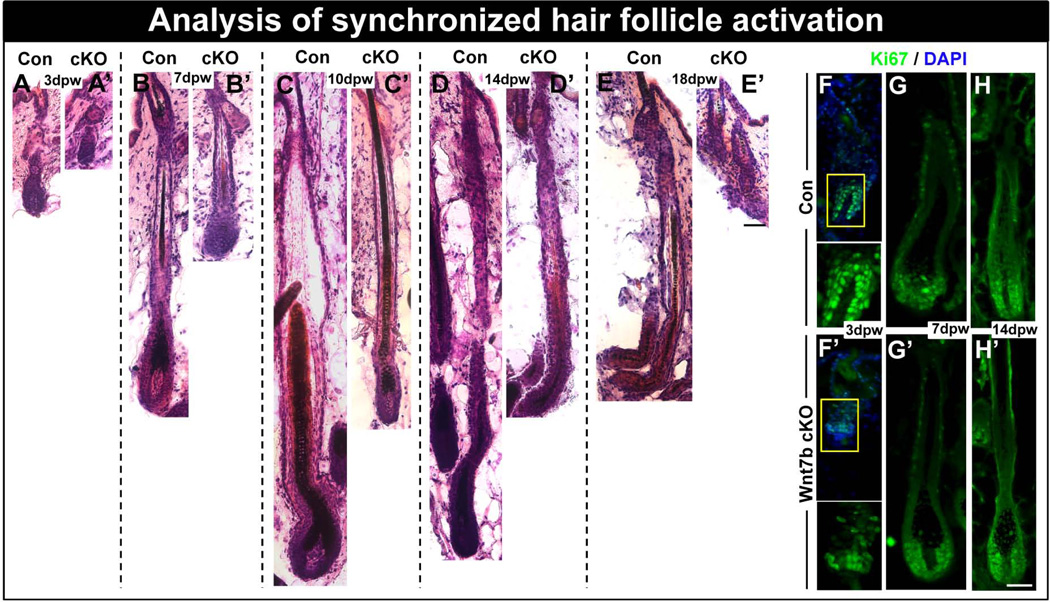
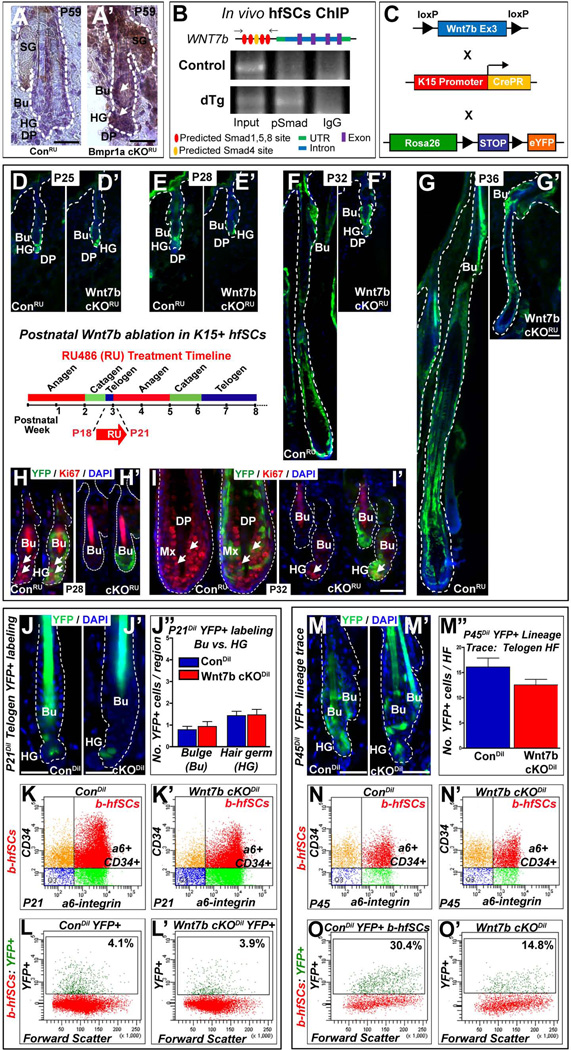
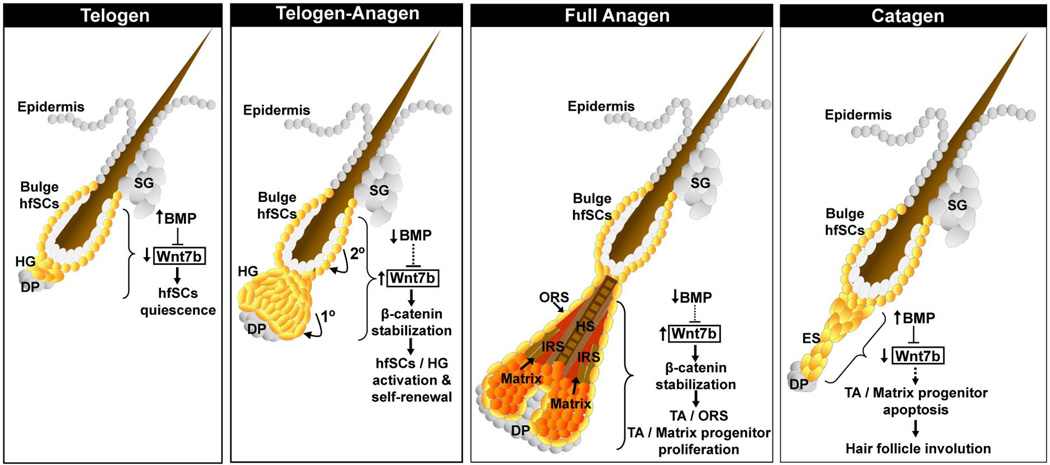
The hair follicle (HF) is an exceptional mini-organ to study the mechanisms which regulate HF morphogenesis, cycling, hair follicle stem cell (hfSCs) homeostasis, and progeny differentiation. During morphogenesis, Wnt signaling is well-characterized in the initiation of HF patterning but less is known about which particular Wnt ligands are required and whether individual Wnt ligands act in an indispensable or redundant manner during postnatal hfSCs anagen onset and HF cycle progression. Previously, we described the function of the bone morphogenetic protein (BMP) signaling target gene WNT7a in intrinsic regulation of hfSCs homeostasis in vivo. Here, we investigated the role of Wnt7b, which was also intrinsically upregulated in hfSCs during physiological and precocious anagen after BMP inhibition in vivo. We demonstrated Wnt7b to be a direct target of canonical BMP signaling in hfSCs and using Wnt7b conditional gene targeting during HF morphogenesis revealed disrupted HF cycling including a shorter anagen, premature catagen onset with overall shorter hair production, and diminished HF differentiation marker expression. Additionally, we observed that postnatal ablation of Wnt7b resulted in delayed HF activation, affecting both the hair germ and bulge hfSCs but still maintaining a two-step sequence of HF stimulation. Interestingly, Wnt7b cKO hfSCs participated in reformation of the new HF bulge, but with slower self-renewal. These findings demonstrate the importance of intrinsic Wnt7b expression in hfSCs regulation and normal HF cycling and surprisingly reveal a nonredundant role for Wnt7b in the control of HF anagen length and catagen entry which was not compensated by other Wnt ligands.
Keywords: Hair follicle stem cells, WNT signaling, Wnt7b.
© AlphaMed Press.
Figures
References
-
- Hardy MH. The secret life of the hair follicle. Trends Genet. 1992;8:55–61. - PubMed
-
- Millar SE. Molecular mechanisms regulating hair follicle development. J Invest Dermatol. 2002;118:216–225. - PubMed
-
- Sengel P, Mauger A. Peridermal cell patterning in the feather-forming skin of the chick embryo. Dev Biol. 1976;51:166–171. - PubMed
-
- Muller-Rover S, Peters EJ, Botchkarev VA, et al. Distinct patterns of NCAM expression are associated with defined stages of murine hair follicle morphogenesis and regression. J Histochem Cytochem. 1998;46:1401–1410. - PubMed
-
- Blanpain C, Lowry WE, Geoghegan A, et al. Self-renewal, multipotency, and the existence of two cell populations within an epithelial stem cell niche. Cell. 2004;118:635–648. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
