

Non-genomic estrogen regulation of ion transport and airway surface liquid dynamics in cystic fibrosis bronchial epithelium
- PMID: 24223826
- PMCID: PMC3817220
- DOI: 10.1371/journal.pone.0078593
Non-genomic estrogen regulation of ion transport and airway surface liquid dynamics in cystic fibrosis bronchial epithelium
Abstract
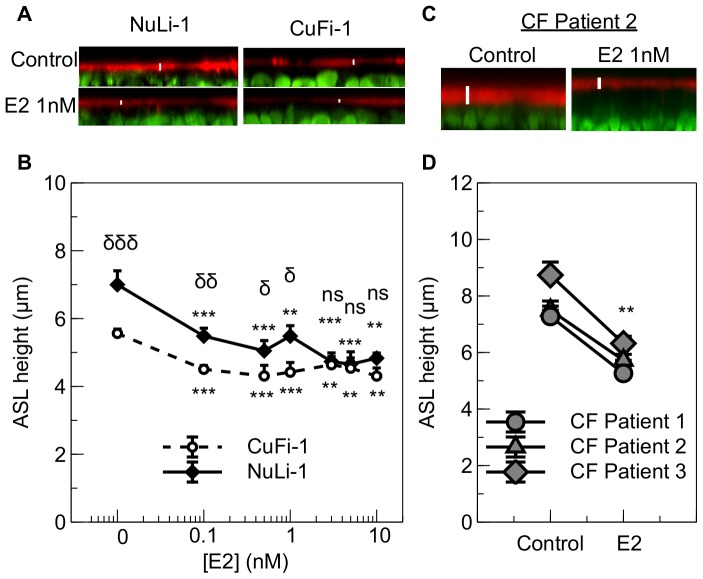
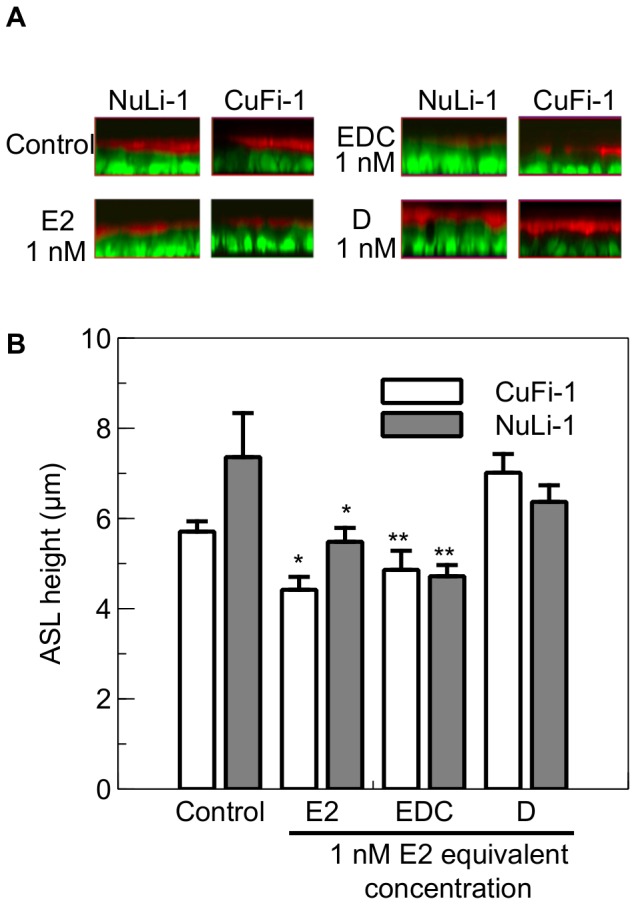
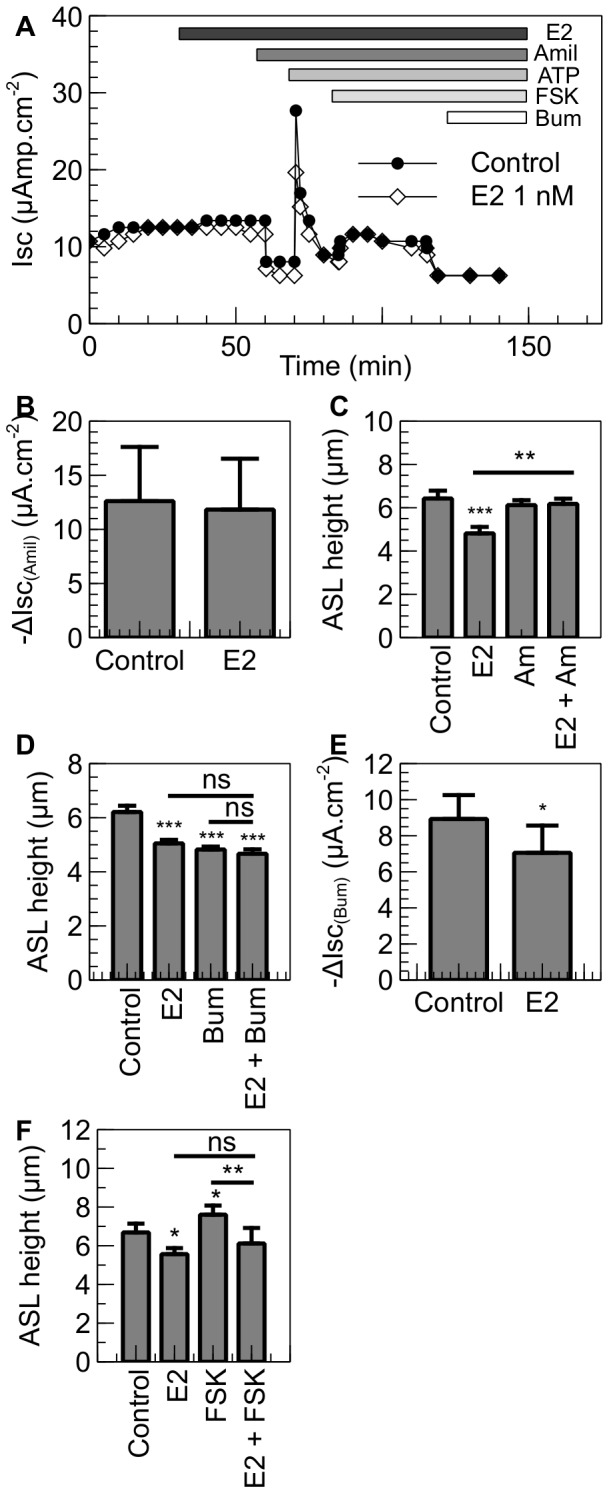
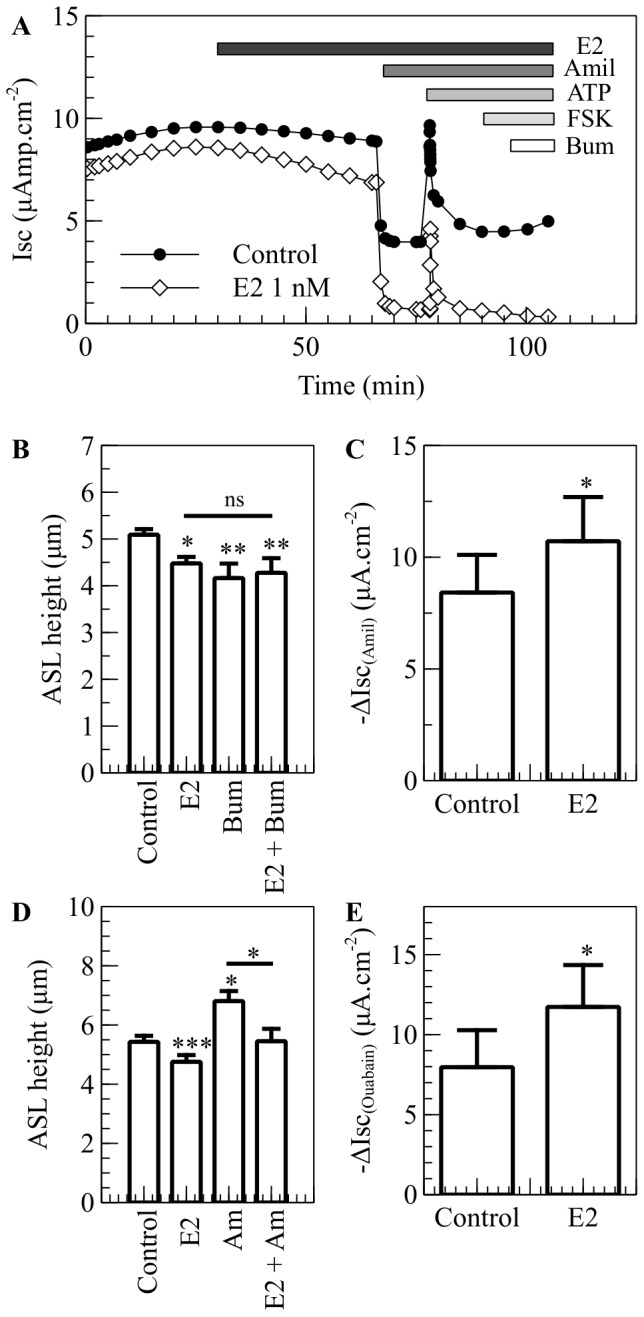
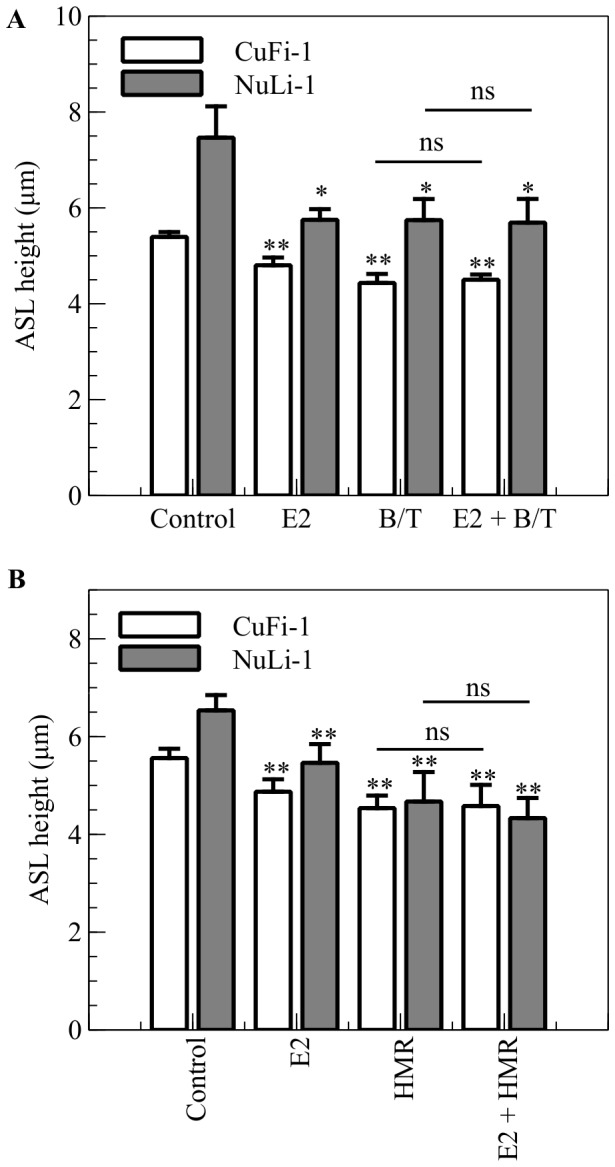
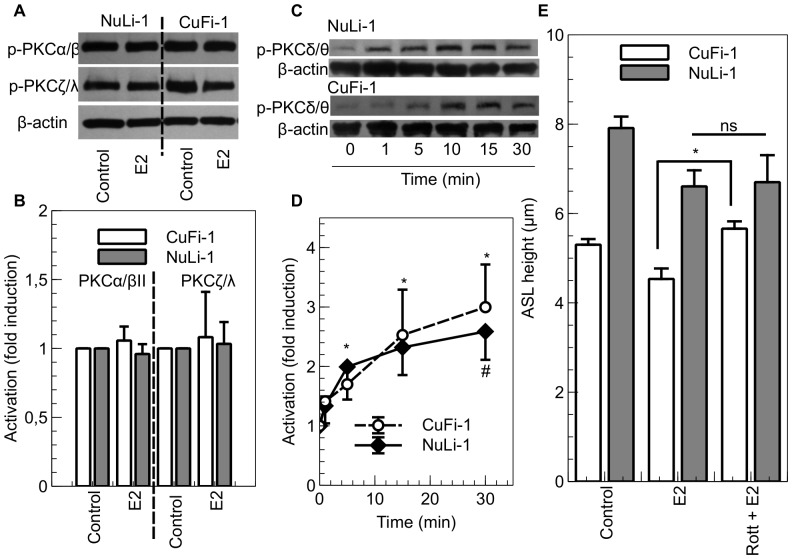
Male cystic fibrosis (CF) patients survive longer than females and lung exacerbations in CF females vary during the estrous cycle. Estrogen has been reported to reduce the height of the airway surface liquid (ASL) in female CF bronchial epithelium. Here we investigated the effect of 17β-estradiol on the airway surface liquid height and ion transport in normal (NuLi-1) and CF (CuFi-1) bronchial epithelial monolayers. Live cell imaging using confocal microscopy revealed that airway surface liquid height was significantly higher in the non-CF cells compared to the CF cells. 17β-estradiol (0.1-10 nM) reduced the airway surface liquid height in non-CF and CF cells after 30 min treatment. Treatment with the nuclear-impeded Estrogen Dendrimer Conjugate mimicked the effect of free estrogen by reducing significantly the airway surface liquid height in CF and non-CF cells. Inhibition of chloride transport or basolateral potassium recycling decreased the airway surface liquid height and 17β-estradiol had no additive effect in the presence of these ion transporter inhibitors. 17β-estradiol decreased bumetanide-sensitive transepithelial short-circuit current in non-CF cells and prevented the forskolin-induced increase in ASL height. 17β-estradiol stimulated an amiloride-sensitive transepithelial current and increased ouabain-sensitive basolateral short-circuit current in CF cells. 17β-estradiol increased PKCδ activity in CF and non-CF cells. These results demonstrate that estrogen dehydrates CF and non-CF ASL, and these responses to 17β-estradiol are non-genomic rather than involving the classical nuclear estrogen receptor pathway. 17β-estradiol acts on the airway surface liquid by inhibiting cAMP-mediated chloride secretion in non-CF cells and increasing sodium absorption via the stimulation of PKCδ, ENaC and the Na(+)/K(+)ATPase in CF cells.
Conflict of interest statement
Figures
Similar articles
-
Ursodeoxycholic acid inhibits ENaC and Na/K pump activity to restore airway surface liquid height in cystic fibrosis bronchial epithelial cells.Steroids. 2019 Nov;151:108461. doi: 10.1016/j.steroids.2019.108461. Epub 2019 Jul 22. Steroids. 2019. PMID: 31344409
-
Soluble mediators, not cilia, determine airway surface liquid volume in normal and cystic fibrosis superficial airway epithelia.J Gen Physiol. 2006 May;127(5):591-604. doi: 10.1085/jgp.200509468. J Gen Physiol. 2006. PMID: 16636206 Free PMC article.
-
Activation of P2RY11 and ATP release by lipoxin A4 restores the airway surface liquid layer and epithelial repair in cystic fibrosis.Am J Respir Cell Mol Biol. 2014 Aug;51(2):178-90. doi: 10.1165/rcmb.2012-0424OC. Am J Respir Cell Mol Biol. 2014. PMID: 24588705
-
Ion and liquid transport across the bronchiolar epithelium.Respir Physiol Neurobiol. 2007 Dec 15;159(3):278-82. doi: 10.1016/j.resp.2007.03.007. Epub 2007 Mar 12. Respir Physiol Neurobiol. 2007. PMID: 17433793 Review.
-
Liquid movement across the surface epithelium of large airways.Respir Physiol Neurobiol. 2007 Dec 15;159(3):256-70. doi: 10.1016/j.resp.2007.06.005. Epub 2007 Jun 17. Respir Physiol Neurobiol. 2007. PMID: 17692578 Free PMC article. Review.
Cited by
-
Inhibition of the sodium-dependent HCO3- transporter SLC4A4, produces a cystic fibrosis-like airway disease phenotype.Elife. 2022 May 30;11:e75871. doi: 10.7554/eLife.75871. Elife. 2022. PMID: 35635440 Free PMC article.
-
Evolution and Prognostic Variables of Cystic Fibrosis in Children and Young Adults: A Narrative Review.Diagnostics (Basel). 2025 Aug 2;15(15):1940. doi: 10.3390/diagnostics15151940. Diagnostics (Basel). 2025. PMID: 40804904 Free PMC article. Review.
-
Changes of androgen and corticosterone metabolites excretion and conversion in cystic fibrosis.Front Endocrinol (Lausanne). 2023 Aug 29;14:1244127. doi: 10.3389/fendo.2023.1244127. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37711888 Free PMC article.
-
Gender differences in bronchiectasis: a real issue?Breathe (Sheff). 2018 Jun;14(2):108-121. doi: 10.1183/20734735.000218. Breathe (Sheff). 2018. PMID: 29875830 Free PMC article. Review.
-
Subcutaneous implant with etonogestrel (Implanon®) for catamenial exacerbations in a patient with cystic fibrosis: a case report.BMC Pulm Med. 2014 Oct 24;14:165. doi: 10.1186/1471-2466-14-165. BMC Pulm Med. 2014. PMID: 25344209 Free PMC article.
References
-
- Beers KW, Thompson MA, Chini EN, Dousa TP (1996) beta-Estradiol inhibits Na+-P(i) cotransport across renal brush border membranes from ovarectomized rats. Biochem Biophys Res Commun 221: 442–445. - PubMed
-
- Hofmeister MV, Damkier HH, Christensen BM, Olde B, Fredrik Leeb-Lundberg LM, et al. (2012) 17beta-Estradiol induces nongenomic effects in renal intercalated cells through G protein-coupled estrogen receptor 1. Am J Physiol Renal Physiol 302: F358–368. - PubMed
-
- Groenestege WM, Hoenderop JG, van den Heuvel L, Knoers N, Bindels RJ (2006) The epithelial Mg2+ channel transient receptor potential melastatin 6 is regulated by dietary Mg2+ content and estrogens. J Am Soc Nephrol 17: 1035–1043. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
