

Body fluid exosomes promote secretion of inflammatory cytokines in monocytic cells via Toll-like receptor signaling
- PMID: 24225954
- PMCID: PMC3868779
- DOI: 10.1074/jbc.M113.512806
Body fluid exosomes promote secretion of inflammatory cytokines in monocytic cells via Toll-like receptor signaling
Abstract
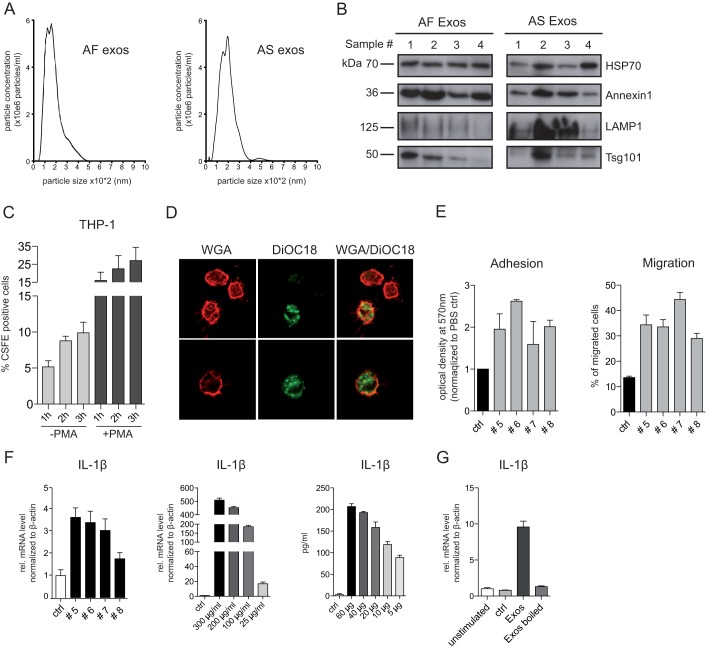
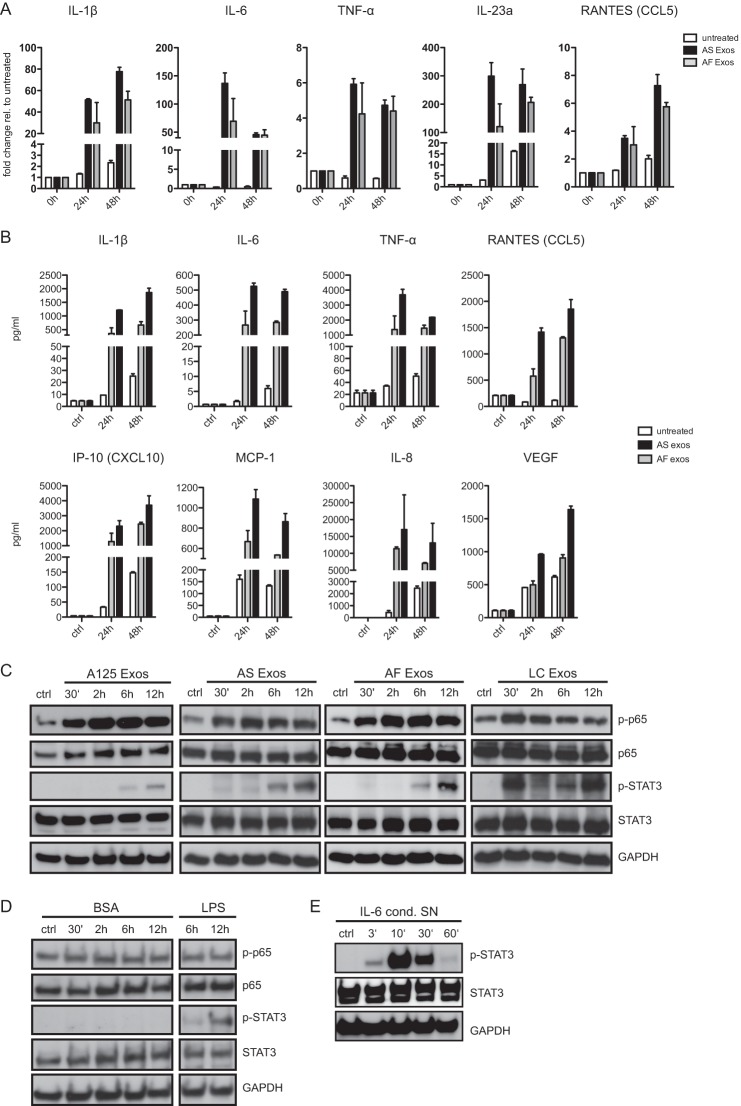
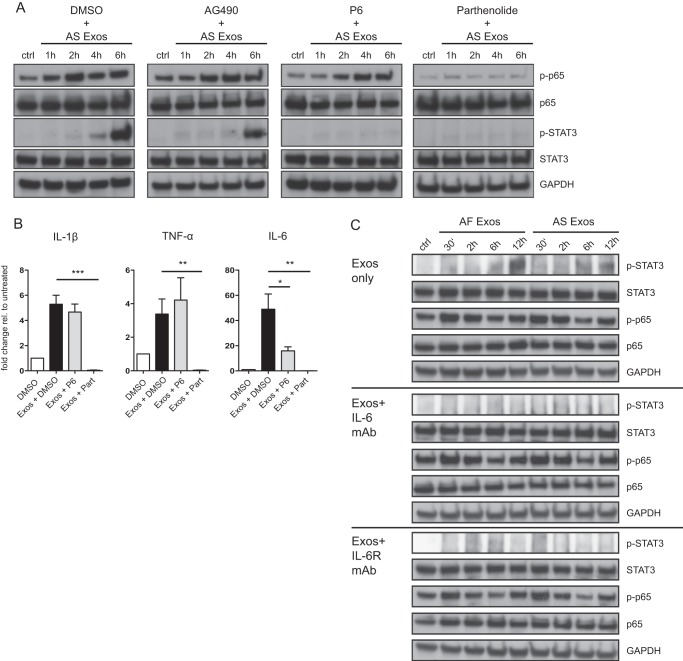
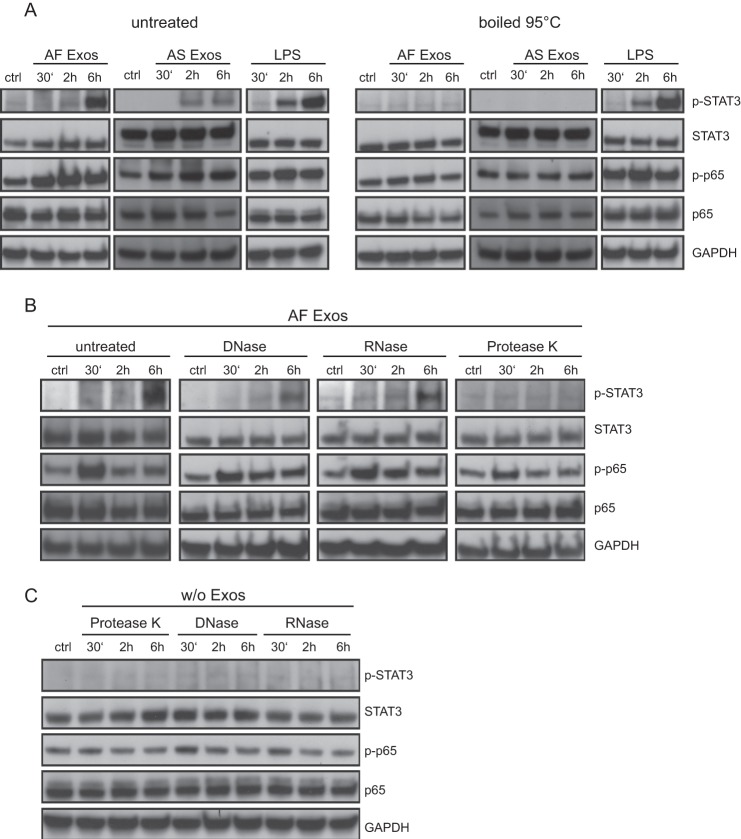
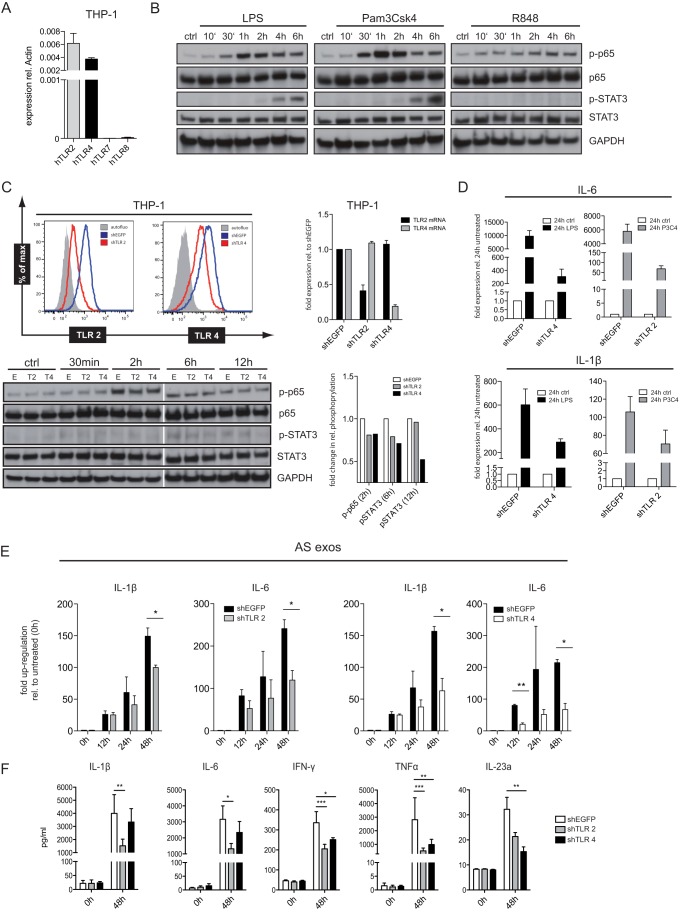
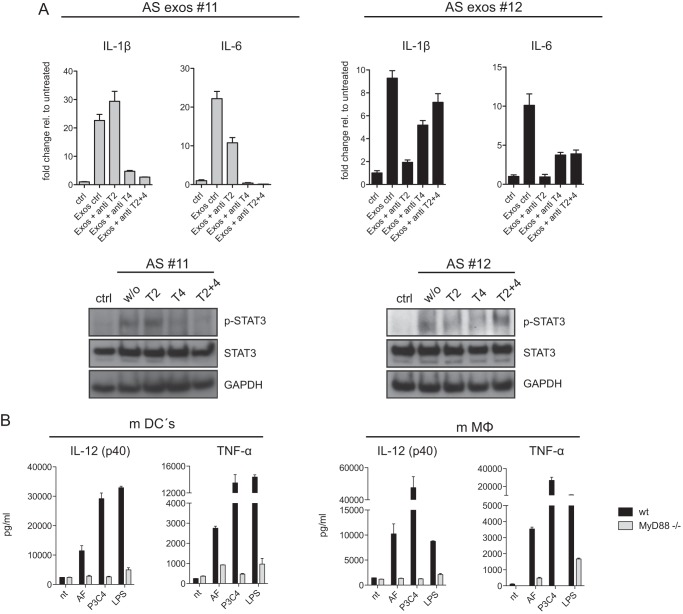
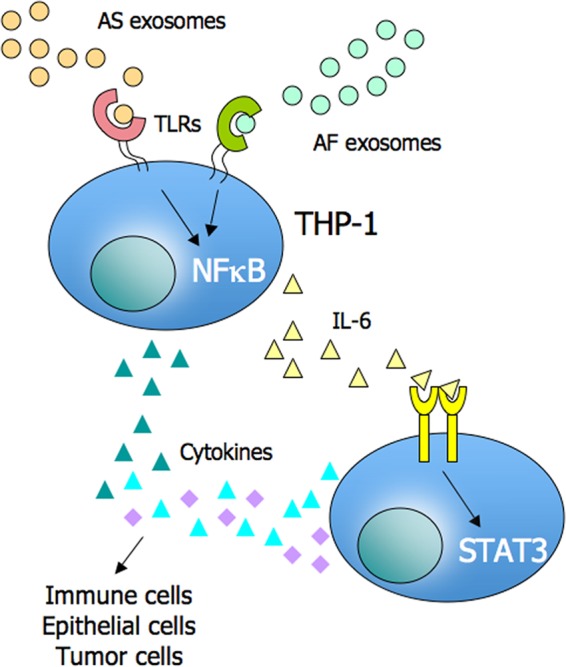
Tumor-derived exosomes have been shown to induce various immunomodulatory effects. However, the underlying signaling pathways are poorly understood. Here, we analyzed the effects of ex vivo-derived exosomes on monocytic cell differentiation/activation using THP-1 cells as model. We isolated exosomes from various body fluids such as amniotic fluid, liver cirrhosis ascites, and malignant ascites of ovarian cancer patients. We observed that exosomes were internalized by THP-1 cells and induced the production of IL-1β, TNF-α, and IL-6. Analysis of the signaling pathways revealed a fast triggering of NFκB and a delayed activation of STAT3. Pharmacologic and antibody-blocking experiments showed that the initial production of IL-6 was instrumental for subsequent activation of STAT3. Importantly, triggering of cell signaling was not a unique property of tumor exosomes but was also observed with exosomes of noncancerous origin. Exosomal signaling was TLR-dependent as the knockdown of Toll-like receptor 2 (TLR2) and TLR4 blocked NFκB and STAT3 activation. Similar results were obtained with TLR-neutralizing antibodies. Exosomes also triggered the release of cytokines from mouse bone marrow-derived dendritic cells or macrophages. This process was MyD88-dependent, further supporting a role of TLR signaling. Our results suggest that exosomes trigger TLR-dependent signaling pathways in monocytic precursor cells but possibly also in other immune cells. This process could be important for the induction of immunosuppressive mechanisms during cancer progression and inflammatory diseases.
Keywords: Amniotic Fluid; Cytokine Induction; Exosomes; Malignant Ascites; NFκB Transcription Factor; STAT3; Toll-like receptor (TLR).
Figures
References
-
- Théry C., Zitvogel L., Amigorena S. (2002) Exosomes: composition, biogenesis and function. Nat. Rev. Immunol. 2, 569–579 - PubMed
-
- Andre F., Schartz N. E., Movassagh M., Flament C., Pautier P., Morice P., Pomel C., Lhomme C., Escudier B., Le Chevalier T., Tursz T., Amigorena S., Raposo G., Angevin E., Zitvogel L. (2002) Malignant effusions and immunogenic tumour-derived exosomes. Lancet 360, 295–305 - PubMed
-
- Hendrix A., Westbroek W., Bracke M., De Wever O. (2010) An ex(o) citing machinery for invasive tumor growth. Cancer Res. 70, 9533–9537 - PubMed
-
- Raiborg C., Rusten T. E., Stenmark H. (2003) Protein sorting into multivesicular endosomes. Curr. Opin. Cell Biol. 15, 446–455 - PubMed
-
- Stoorvogel W., Kleijmeer M. J., Geuze H. J., Raposo G. (2002) The biogenesis and functions of exosomes. Traffic 3, 321–330 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
