

Systems metabolic engineering of Corynebacterium glutamicum for production of the chemical chaperone ectoine
- PMID: 24228689
- PMCID: PMC4225761
- DOI: 10.1186/1475-2859-12-110
Systems metabolic engineering of Corynebacterium glutamicum for production of the chemical chaperone ectoine
Abstract
Background: The stabilizing and function-preserving effects of ectoines have attracted considerable biotechnological interest up to industrial scale processes for their production. These rely on the release of ectoines from high-salinity-cultivated microbial producer cells upon an osmotic down-shock in rather complex processor configurations. There is growing interest in uncoupling the production of ectoines from the typical conditions required for their synthesis, and instead design strains that naturally release ectoines into the medium without the need for osmotic changes, since the use of high-salinity media in the fermentation process imposes notable constraints on the costs, design, and durability of fermenter systems.
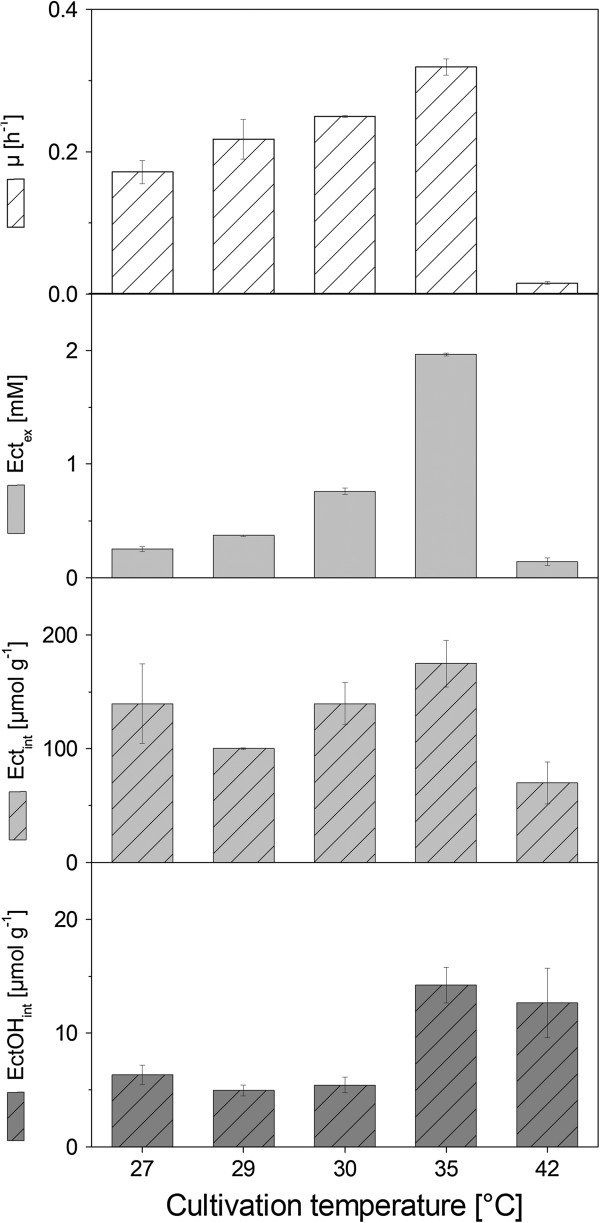
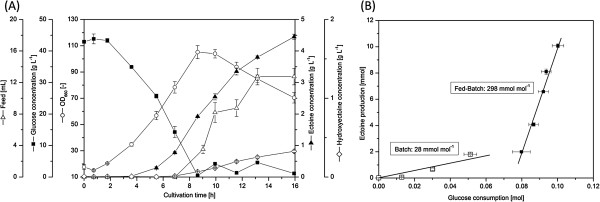
Results: Here, we used a Corynebacterium glutamicum strain as a cellular chassis to establish a microbial cell factory for the biotechnological production of ectoines. The implementation of a mutant aspartokinase enzyme ensured efficient supply of L-aspartate-beta-semialdehyde, the precursor for ectoine biosynthesis. We further engineered the genome of the basic C. glutamicum strain by integrating a codon-optimized synthetic ectABCD gene cluster under expressional control of the strong and constitutive C. glutamicum tuf promoter. The resulting recombinant strain produced ectoine and excreted it into the medium; however, lysine was still found as a by-product. Subsequent inactivation of the L-lysine exporter prevented the undesired excretion of lysine while ectoine was still exported. Using the streamlined cell factory, a fed-batch process was established that allowed the production of ectoine with an overall productivity of 6.7 g L(-1) day(-1) under growth conditions that did not rely on the use of high-salinity media.
Conclusions: The present study describes the construction of a stable microbial cell factory for recombinant production of ectoine. We successfully applied metabolic engineering strategies to optimize its synthetic production in the industrial workhorse C. glutamicum and thereby paved the way for further improvements in ectoine yield and biotechnological process optimization.
Figures


References
-
- Wood JM, Bremer E, Csonka LN, Kraemer R, Poolman B, van der Heide T, Smith LT. Osmosensing and osmoregulatory compatible solute accumulation by bacteria. Comp Biochem Physiol A Mol Integr Physiol. 2001;130:437–460. - PubMed
-
- Kempf B, Bremer E. Uptake and synthesis of compatible solutes as microbial stress responses to high-osmolality environments. Arch Microbiol. 1998;170:319–330. - PubMed
-
- Bremer E, Krämer R. In: Bacterial Stress Responses. Storz G, Hengge-Aronis R, editor. Washington D.C.: ASM Press; 2000. Coping with osmotic challenges: osmoregulation through accumulation and release of compatible solutes; pp. 79–97.
-
- Diamant S, Eliahu N, Rosenthal D, Goloubinoff P. Chemical chaperones regulate molecular chaperones in vitro and in cells under combined salt and heat stresses. J Biol Chem. 2001;276:39586–39591. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
