

Ciliogenesis and cerebrospinal fluid flow in the developing Xenopus brain are regulated by foxj1
- PMID: 24229449
- PMCID: PMC3848805
- DOI: 10.1186/2046-2530-2-12
Ciliogenesis and cerebrospinal fluid flow in the developing Xenopus brain are regulated by foxj1
Abstract
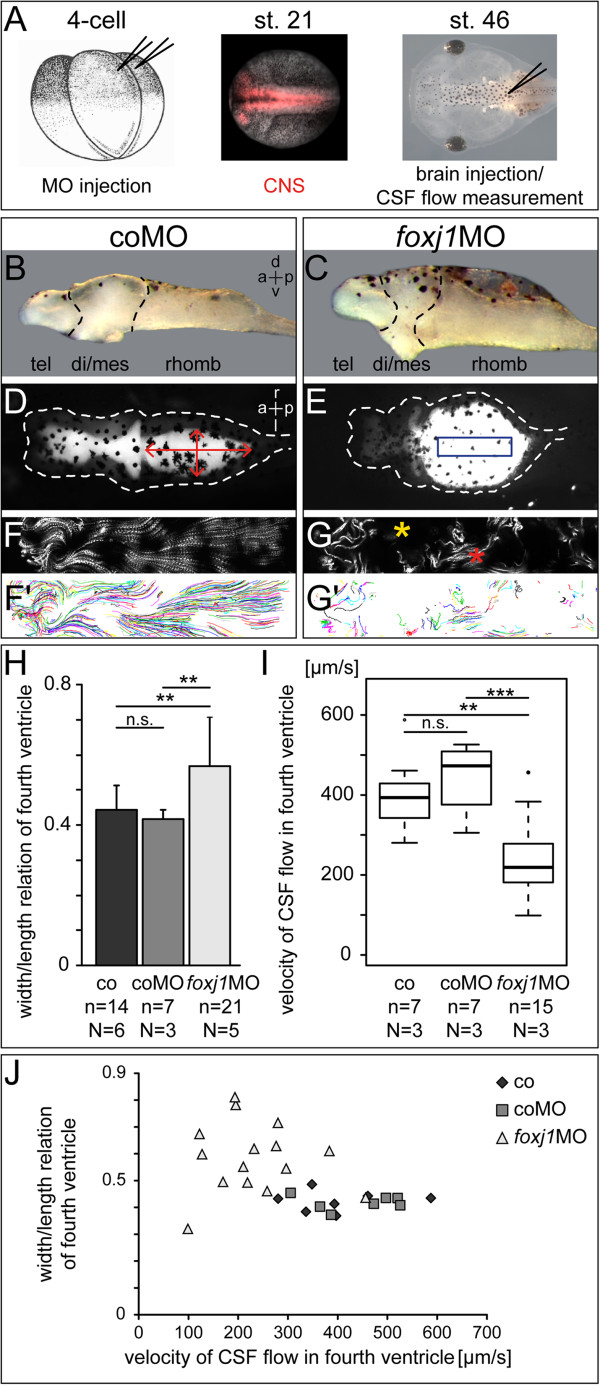
Background: Circulation of cerebrospinal fluid (CSF) through the ventricular system is driven by motile cilia on ependymal cells of the brain. Disturbed ciliary motility induces the formation of hydrocephalus, a pathological accumulation of CSF resulting in ventricle dilatation and increased intracranial pressure. The mechanism by which loss of motile cilia causes hydrocephalus has not been elucidated. The aim of this study was: (1) to provide a detailed account of the development of ciliation in the brain of the African clawed frog Xenopus laevis; and (2) to analyze the relevance of ependymal cilia motility for CSF circulation and brain ventricle morphogenesis in Xenopus.
Methods: Gene expression analysis of foxj1, the bona fide marker for motile cilia, was used to identify potentially ciliated regions in the developing central nervous system (CNS) of the tadpole. Scanning electron microscopy (SEM) was used to reveal the distribution of mono- and multiciliated cells during successive stages of brain morphogenesis, which was functionally assessed by bead injection and video microscopy of ventricular CSF flow. An antisense morpholino oligonucleotide (MO)-mediated gene knock-down that targeted foxj1 in the CNS was applied to assess the role of motile cilia in the ventricles.
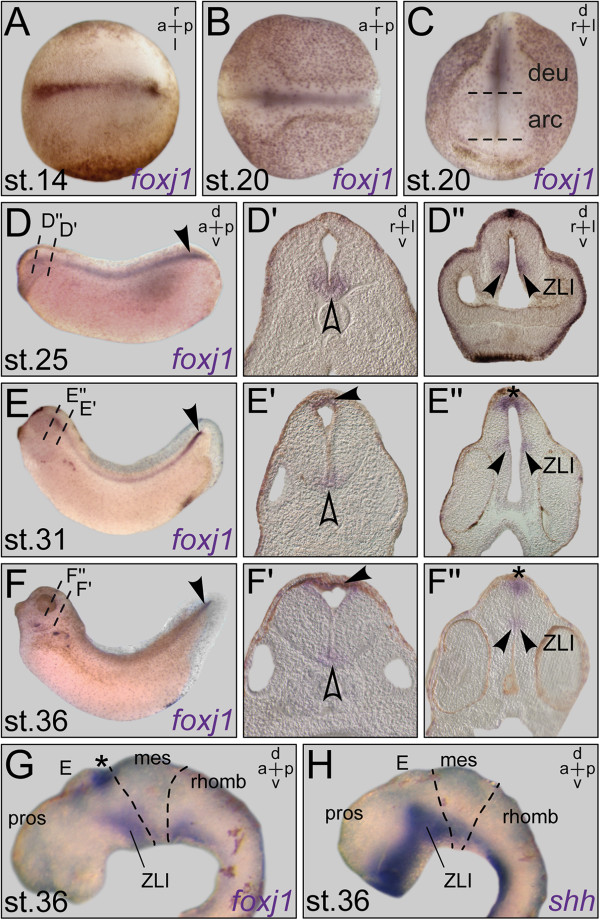
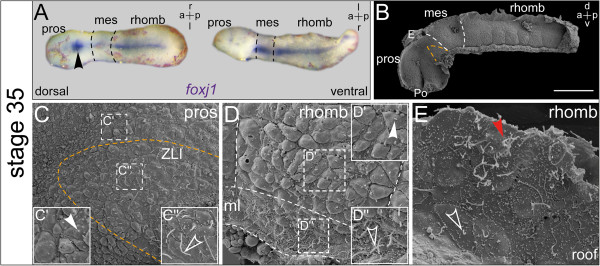
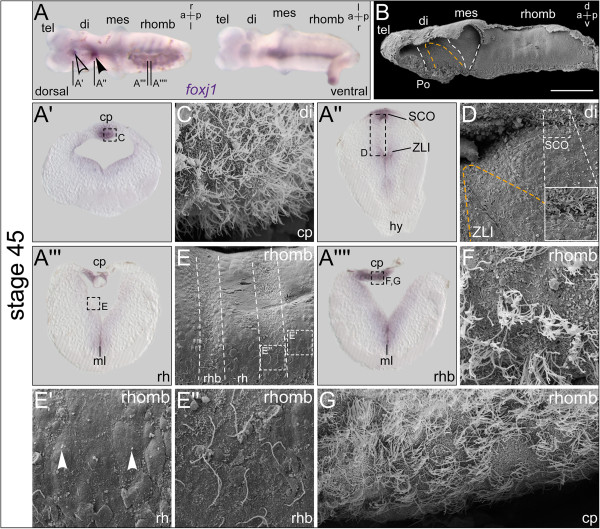
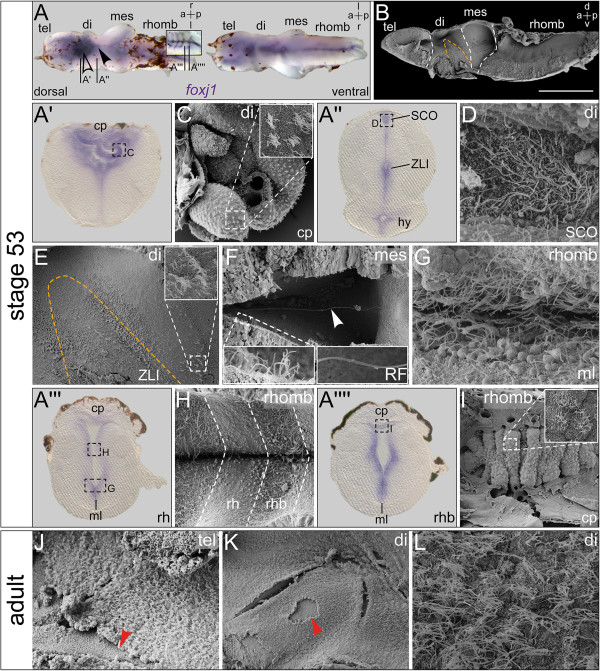
Results: RNA transcripts of foxj1 in the CNS were found from neurula stages onwards. Following neural tube closure, foxj1 expression was seen in distinct ventricular regions such as the zona limitans intrathalamica (ZLI), subcommissural organ (SCO), floor plate, choroid plexus (CP), and rhombomere boundaries. In all areas, expression of foxj1 preceded the outgrowth of monocilia and the subsequent switch to multiciliated ependymal cells. Cilia were absent in foxj1 morphants, causing impaired CSF flow and fourth ventricle hydrocephalus in tadpole-stage embryos.
Conclusions: Motile ependymal cilia are important organelles in the Xenopus CNS, as they are essential for the circulation of CSF and maintenance of homeostatic fluid pressure. The Xenopus CNS ventricles might serve as a novel model system for the analysis of human ciliary genes whose deficiency cause hydrocephalus.
Figures

References
-
- Sawamoto K, Wichterle H, Gonzalez-Perez O, Cholfin J, Yamada M, Spassky N, Murcia N, Garcia-Verdugo J, Marin O, Rubenstein J, Tessier-Lavigne M, Okano H, Alvarez-Buylla A. New neurons follow the flow of cerebrospinal fluid in the adult brain. Science. 2006;311:629–632. doi: 10.1126/science.1119133. - DOI - PubMed
-
- Iba?ez-Tallon I, Pagenstecher A, Fliegauf M, Olbrich H, Kispert A, Ketelsen UP, North A, Heintz N, Omran H. Dysfunction of axonemal dynein heavy chain Mdnah5 inhibits ependymal flow and reveals a novel mechanism for hydrocephalus formation. Hum Mol Genet. 2004;13:2133–2141. doi: 10.1093/hmg/ddh219. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
