

doi: 10.1038/nchembio.1403.
Adaptive immune activation: glycosylation does matter
Affiliations
- PMID: 24231619
- PMCID: PMC3966069
- DOI: 10.1038/nchembio.1403
Item in Clipboard
Adaptive immune activation: glycosylation does matter
Nat Chem Biol.
2013 Dec.
Abstract
Major histocompatibility complex (MHC) class I and II are glycoproteins that can present antigenic peptides at the cell surface for recognition and activation of circulating T lymphocytes. Here, the importance of the modification of protein antigens by glycans on cellular uptake, proteolytic processing, presentation by MHC and subsequent T-cell priming is reviewed. Antigen glycosylation is important for a number of diseases and vaccine design. All of the key proteins involved in antigen recognition and the orchestration of downstream effector functions are glycosylated. The influence of protein glycosylation on immune function and disease is covered.
Figures
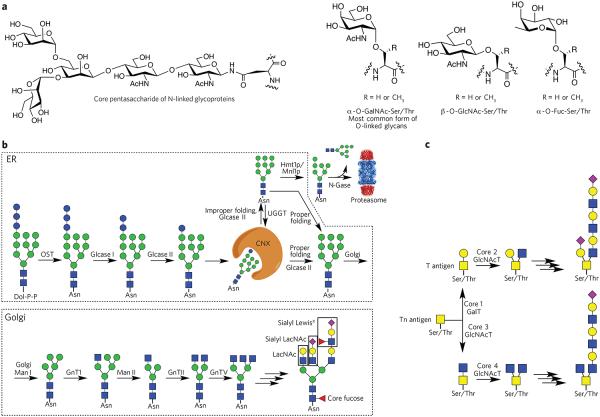
(a) Common types of protein glycosylation: core pentasaccharide common to all N-linked glycans and various types of O-glycosylation. (b) Biosynthesis of N-glycans. The biosynthesis of N-glycans is initiated in the ER by the transfer of a dolichol (Dol-P-P)-linked glycan to an asparagine moiety of an Asn-Xaa-Ser/Thr sequon of a polypeptide. In the ER, protein glycosylation functions in the quality control of protein biosynthesis. Calnexin (CNX) is a chaperone that keeps the unfolded protein in the ER. Improperly folded glycans can encounter the α-mannosidase MNS1, which removes a terminal mannoside, providing a glycoform that will be transported to the cytosol. Here, an N-glycanase (N-Gase) removes the N-linked glycans, and the resulting protein is then imported into the proteasome for proteolysis. Properly folded proteins are transported to the Golgi, where an array of glycosyltransferases diversifies the various antennae of the glycans to give an array of complex structures. The resulting complex-type glycoproteins are transported to the cell surface or secreted. OST, oligosaccharyltransferase; glcase, α-glucosidase; UGGT, UDP–glucose-glycoprotein glucosyltransferase; Hmt1p, HnRNP methyltransferase 1; Mnl1p, mannosidase-like protein 1; N-gase, N-glucosidase; Man, α-mannosidase; GnT, N-acetyl glucosaminyltransferase. (c) Biosynthesis of O-glycans. The biosynthesis of O-glycans that start the structure with an O-GalNAc moiety begins in the Golgi, where various polypeptide polypeptide GalNAc transferases (GlcNAcT) attach a GalNAc residue. The resulting glycoprotein can be converted into various core structures that can be diversified by a range of glycosyltransferases.
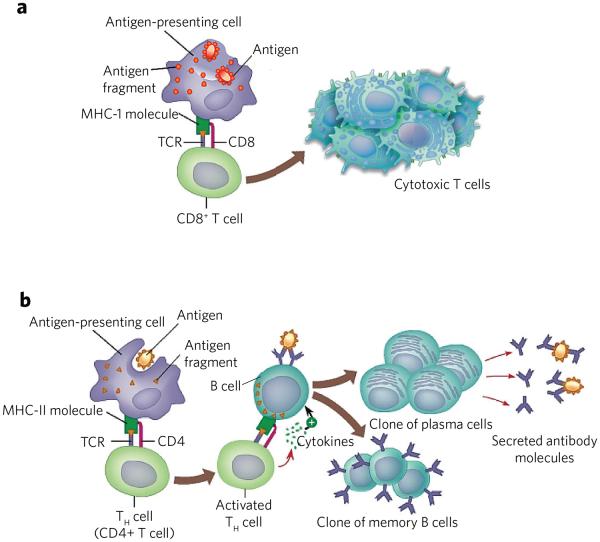
(a) MHC-I presents peptides derived from endogenous antigens. Antigen is proteolyzed by the immune proteasome and specific peptides loaded to MHC-I. The resulting MHC-I–peptide complex is transported to the cell surface, where it can be recognized by the TCR of CD8+ cells. The resulting CD8+ cells proliferate to give cytotoxic T cells. These cells can kill other cells that present the same antigen in complex with MHC-I. (b) Endocytosis of antigen by antigen-presenting cells followed by proteolysis by lysosomes provides peptides that can be loaded on MHC-II. The resulting complex is transported to the cell surface where it can activate CD4+ cells. The resulting cells can activate B cells that present the same antigenic peptide in complex with MHC-II. The activated B cell differentiates into plasma cells that secrete antibodies and memory B cells.
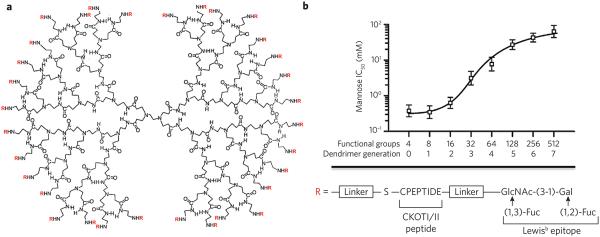
(a) Glycopeptide dendrimers were synthesized by attaching a glycopeptide to the primary amines of PAMAM dendrimers by functionalization with a maleimide-containing linker followed by reaction with the thiol of the C-terminal cysteine of the CKOTI/II peptide, which was further extended by a Lewisb epitope. The structure of a third-generation dendrimer is shown. (b) Cellular binding of glycopeptide dendrimers. The glycopeptide dendrimers were labeled with a fluorophore and incubated with mutant DC-SIGN–expressing cells in the presence of increasing concentrations of mannose as a binding inhibitor. The resulting half-maximum inhibitory concentration (IC50) values are plotted against the number of functional groups attached to the dendrimer. Reprinted with permission from reference 52.
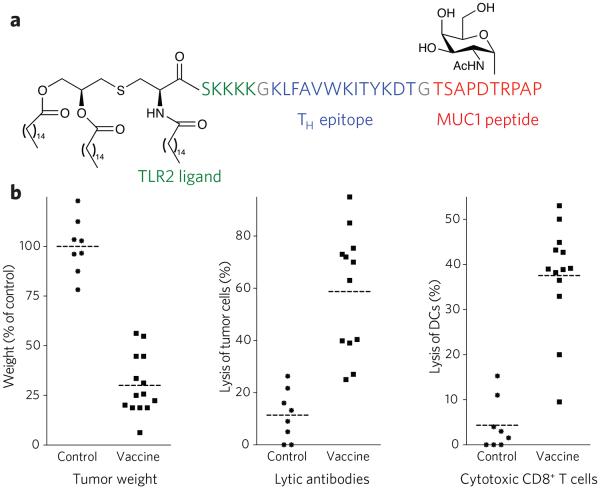
(a) Chemical structure of synthetic vaccine. (b) Left, glycosylated multi-component vaccine reduces MMT tumor burden in MUC1.Tg mice. MUC1.Tg mice were immunized with empty liposomes (EL) as control or with liposomes containing the vaccine. Three biweekly immunizations were given before a tumor challenge with MUC1-expressing MMT tumor cells followed by one boost one week after. Middle, induction of antibody-dependent cell-mediated cytotoxicity (ADCC). Right, induction of CD8+ cytolytic T cells in MUC1.Tg mice. Reprinted with permission from reference 79. Each data point represents an individual mouse, and the horizontal lines indicate the mean for the group of mice.
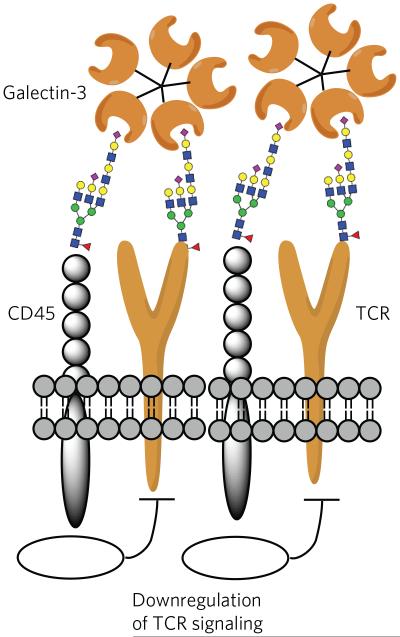
Galectin-3 can organize the TCR and CD45 of T cells into lattices by binding their glycans. In this way, galectin-3 is proposed to block direct interactions of TCRs. Furthermore, the phosphatase activity of CD45 causes downregulation of T-cell signaling.
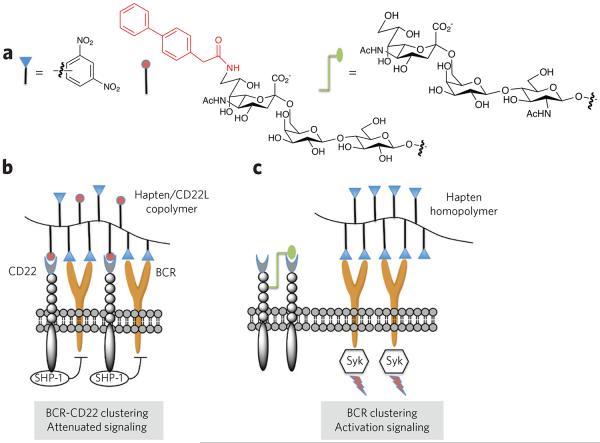
(a) Polymers were designed that contain dinitrophenyl as a hapten, a carbohydrate ligand for CD22 (CD22L) or both. A biphenyl moiety at C6 of the sialoside of the CD22L greatly increases the affinity of binding. (b) The copolymer bearing the hapten and CD22 ligand induces clustering of the B-cell receptor (BCR) and CD22, which results in a suppression of B-cell signaling and tolerance. In this case, the immune-receptor tyrosine-based inhibitory motifs of CD22 recruit the inhibitory phosphatases SHP-1 and SHIP, which suppress BCR signaling through unknown mechanisms. It is thought that trans interactions with 2,6-linked sialosides of self cells induce tolerance through a similar mechanism. (c) A polymer containing only hapten mimics a foreign entity that can cluster BCRs, resulting in cell signaling. In this case, CD22 is largely excluded from the BCR signaling complex. CD22 is a glycoprotein, and it has been shown that it can bind with its own glycans forming homomultimeric protein complexes.
References
-
- Roth J. Protein N-glycosylation along the secretory pathway: relationship to organelle topography and function, protein quality control, and cell interactions. Chem. Rev. 2002;102:285–303. - PubMed
-
- Kleene R, Schachner M. Glycans and neural cell interactions. Nat. Rev. Neurosci. 2004;5:195–208. - PubMed
-
- Ohtsubo K, Marth JD. Glycosylation in cellular mechanisms of health and disease. Cell. 2006;126:855–867. - PubMed
-
- Prescher JA, Bertozzi CR. Chemical technologies for probing glycans. Cell. 2006;126:851–854. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
