

Building and remodelling Cullin-RING E3 ubiquitin ligases
- PMID: 24232186
- PMCID: PMC3849489
- DOI: 10.1038/embor.2013.173
Building and remodelling Cullin-RING E3 ubiquitin ligases
Abstract
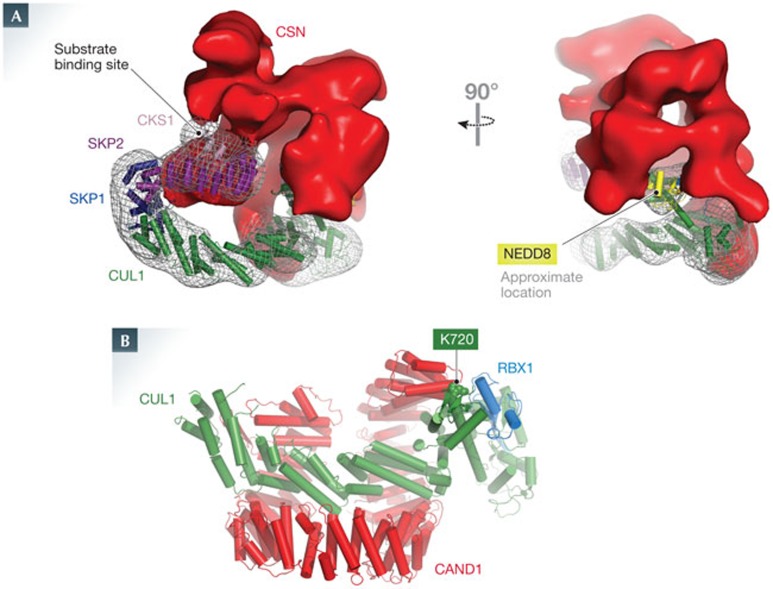
Cullin-RING E3 ubiquitin ligases (CRLs) control a plethora of biological pathways through targeted ubiquitylation of signalling proteins. These modular assemblies use substrate receptor modules to recruit specific targets. Recent efforts have focused on understanding the mechanisms that control the activity state of CRLs through dynamic alterations in CRL architecture. Central to these processes are cycles of cullin neddylation and deneddylation, as well as exchange of substrate receptor modules to re-sculpt the CRL landscape, thereby responding to the cellular requirements to turn over distinct proteins in different contexts. This review is focused on how CRLs are dynamically controlled with an emphasis on how cullin neddylation cycles are integrated with receptor exchange.
Conflict of interest statement
J.W.H. is a consultant for Millenium Pharmaceutics.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
