

Transcriptome signatures of class I and III stress response deregulation in Lactobacillus plantarum reveal pleiotropic adaptation
- PMID: 24238744
- PMCID: PMC3842655
- DOI: 10.1186/1475-2859-12-112
Transcriptome signatures of class I and III stress response deregulation in Lactobacillus plantarum reveal pleiotropic adaptation
Abstract
Background: To cope with environmental challenges bacteria possess sophisticated defense mechanisms that involve stress-induced adaptive responses. The canonical stress regulators CtsR and HrcA play a central role in the adaptations to a plethora of stresses in a variety of organisms. Here, we determined the CtsR and HrcA regulons of the lactic acid bacterium Lactobacillus plantarum WCFS1 grown under reference (28°C) and elevated (40°C) temperatures, using ctsR, hrcA, and ctsR-hrcA deletion mutants.
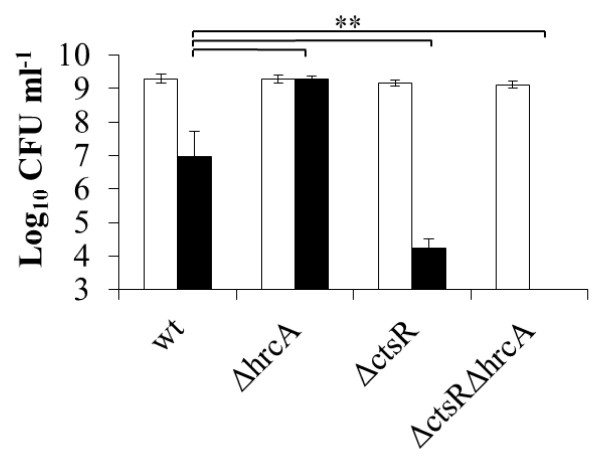
Results: While the maximum specific growth rates of the mutants and the parental strain were similar at both temperatures (0.33 ± 0.02 h(-1) and 0.34 ± 0.03 h(-1), respectively), DNA microarray analyses revealed that the CtsR or HrcA deficient strains displayed altered transcription patterns of genes encoding functions involved in transport and binding of sugars and other compounds, primary metabolism, transcription regulation, capsular polysaccharide biosynthesis, as well as fatty acid metabolism. These transcriptional signatures enabled the refinement of the gene repertoire that is directly or indirectly controlled by CtsR and HrcA of L. plantarum. Deletion of both regulators, elicited transcriptional changes of a large variety of additional genes in a temperature-dependent manner, including genes encoding functions involved in cell-envelope remodeling. Moreover, phenotypic assays revealed that both transcription regulators contribute to regulation of resistance to hydrogen peroxide stress. The integration of these results allowed the reconstruction of CtsR and HrcA regulatory networks in L. plantarum, highlighting the significant intertwinement of class I and III stress regulons.
Conclusions: Taken together, our results enabled the refinement of the CtsR and HrcA regulatory networks in L. plantarum, illustrating the complex nature of adaptive stress responses in this bacterium.
Figures





References
-
- Lee NK, Yun CW, Kim SW, Chang HI, Kang CW, Paik HD. Screening of Lactobacilli derived from chicken feces and partial characterization of Lactobacillus acidophilus A12 as an animal probiotics. J Microbiol Biotechnol. 2008;18(2):338–342. - PubMed
-
- Siezen RJ, Tzeneva VA, Castioni A, Wels M, Phan HT, Rademaker JL, Starrenburg MJ, Kleerebezem M, Molenaar D, Van Hylckama Vlieg JE. Phenotypic and genomic diversity of Lactobacillus plantarum strains isolated from various environmental niches. Environ Microbiol. 2010;12(3):758–773. doi: 10.1111/j.1462-2920.2009.02119.x. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
