

Dissecting local circuits: parvalbumin interneurons underlie broad feedback control of olfactory bulb output
- PMID: 24239125
- PMCID: PMC3932159
- DOI: 10.1016/j.neuron.2013.08.027
Dissecting local circuits: parvalbumin interneurons underlie broad feedback control of olfactory bulb output
Abstract
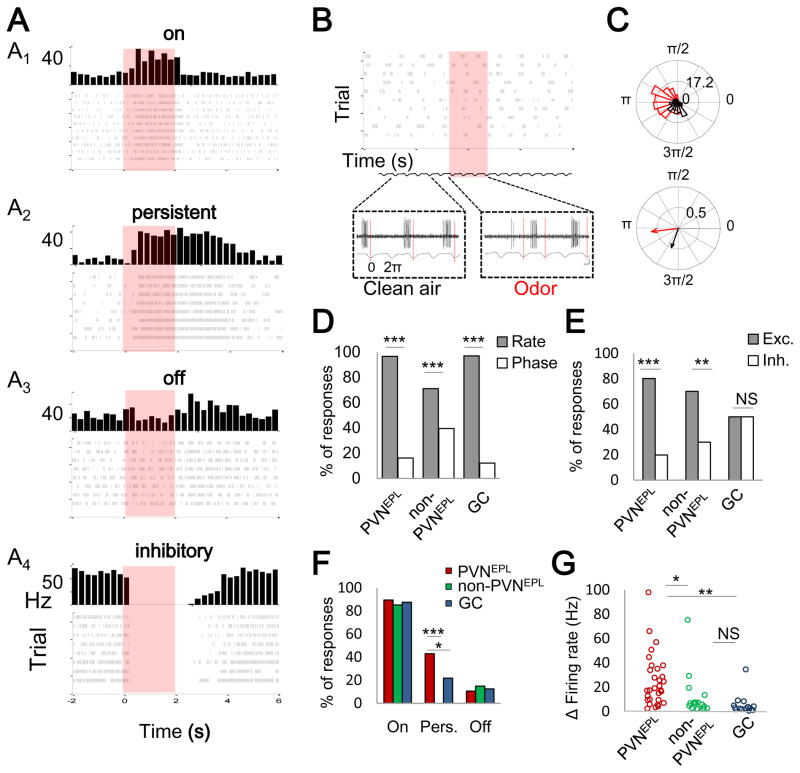
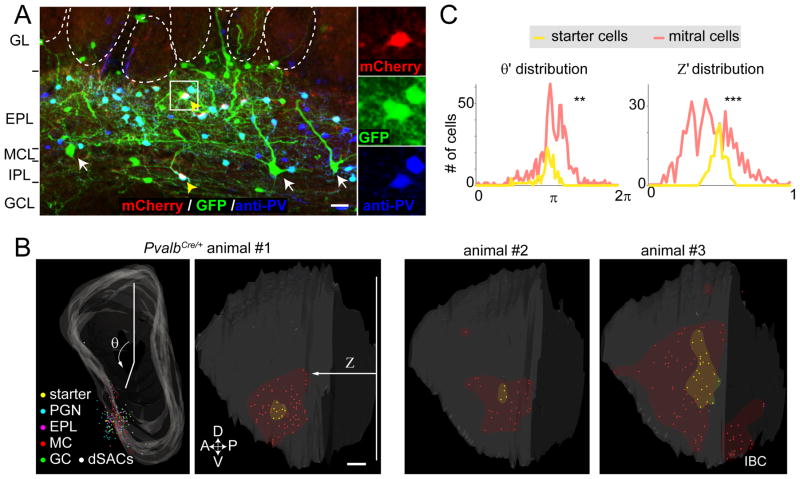
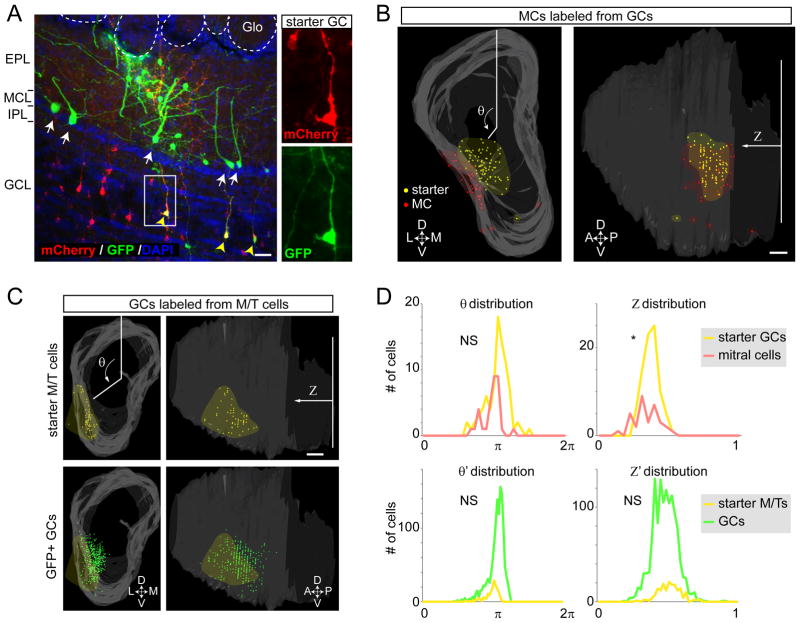
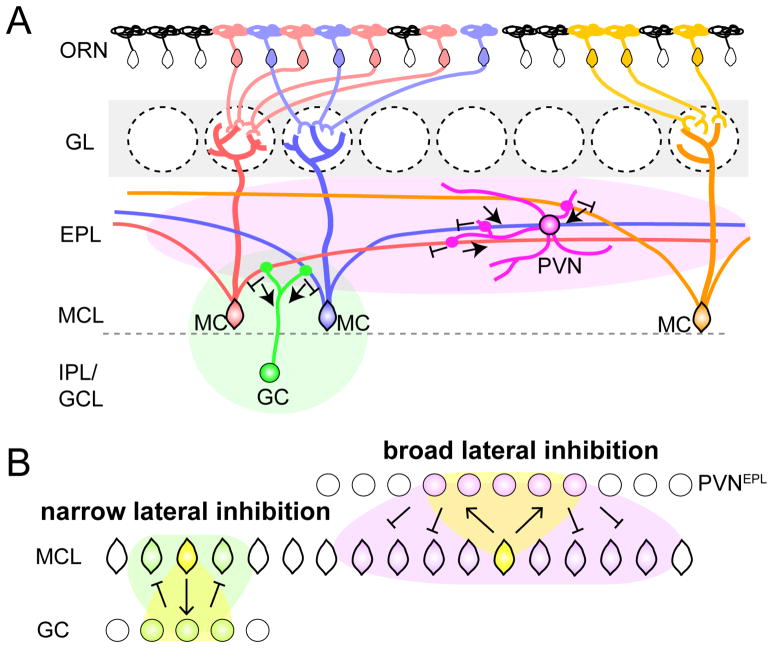
In the mouse olfactory bulb, information from sensory neurons is extensively processed by local interneurons before being transmitted to the olfactory cortex by mitral and tufted (M/T) cells. The precise function of these local networks remains elusive because of the vast heterogeneity of interneurons, their diverse physiological properties, and their complex synaptic connectivity. Here we identified the parvalbumin interneurons (PVNs) as a prominent component of the M/T presynaptic landscape by using an improved rabies-based transsynaptic tracing method for local circuits. In vivo two-photon-targeted patch recording revealed that PVNs have exceptionally broad olfactory receptive fields and exhibit largely excitatory and persistent odor responses. Transsynaptic tracing indicated that PVNs receive direct input from widely distributed M/T cells. Both the anatomical and functional extent of this M/T→PVN→M/T circuit contrasts with the narrowly confined M/T→granule cell→M/T circuit, suggesting that olfactory information is processed by multiple local circuits operating at distinct spatial scales.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
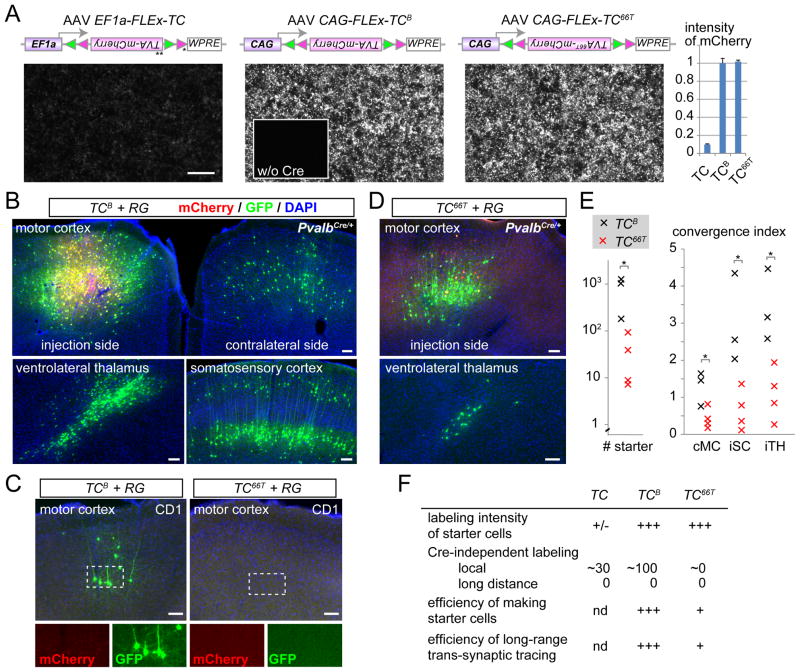
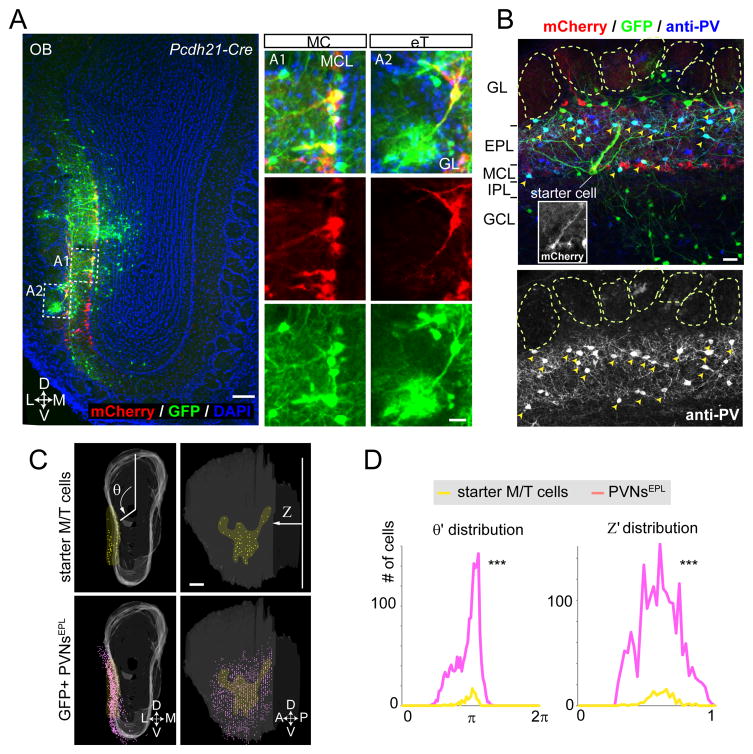
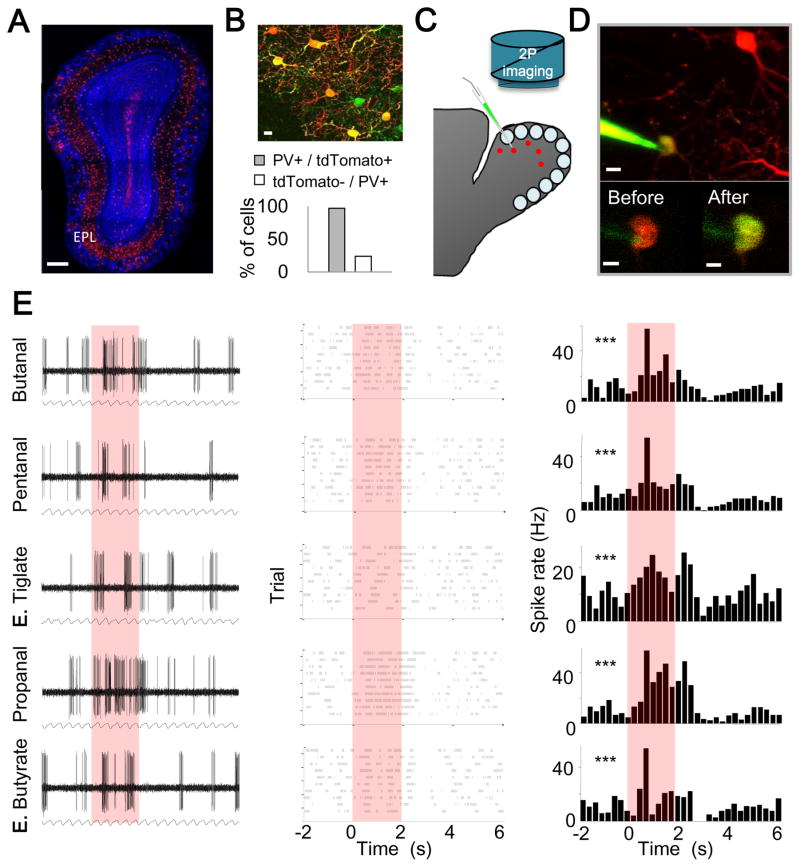
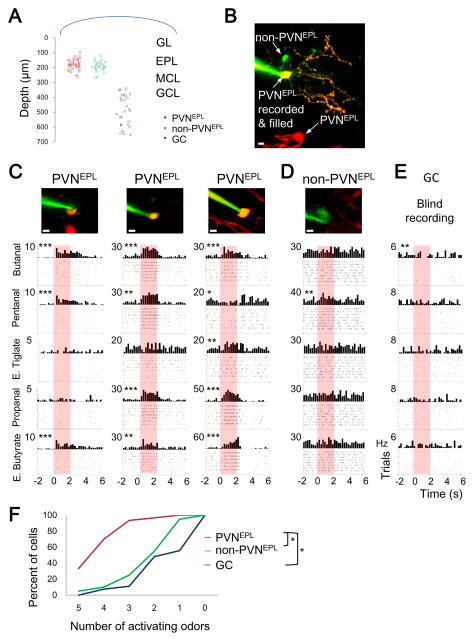
Comment in
-
Sensory systems: connecting olfaction.Nat Rev Neurosci. 2014 Jan;15(1):4-5. doi: 10.1038/nrn3652. Epub 2013 Dec 4. Nat Rev Neurosci. 2014. PMID: 24301068 No abstract available.
-
Division of labor for division: inhibitory interneurons with different spatial landscapes in the olfactory system.Neuron. 2013 Dec 4;80(5):1106-9. doi: 10.1016/j.neuron.2013.11.013. Neuron. 2013. PMID: 24314722 Free PMC article.
References
-
- Aungst JL, Heyward PM, Puche AC, Karnup SV, Hayar A, Szabo G, Shipley MT. Centre-surround inhibition among olfactory bulb glomeruli. Nature. 2003;426:623–629. - PubMed
-
- Cruikshank SJ, Lewis TJ, Connors BW. Synaptic basis for intense thalamocortical activation of feedforward inhibitory cells in neocortex. Nat Neurosci. 2007;10:462–468. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
