

A bacterial toxin inhibits DNA replication elongation through a direct interaction with the β sliding clamp
- PMID: 24239291
- PMCID: PMC3918436
- DOI: 10.1016/j.molcel.2013.10.014
A bacterial toxin inhibits DNA replication elongation through a direct interaction with the β sliding clamp
Abstract
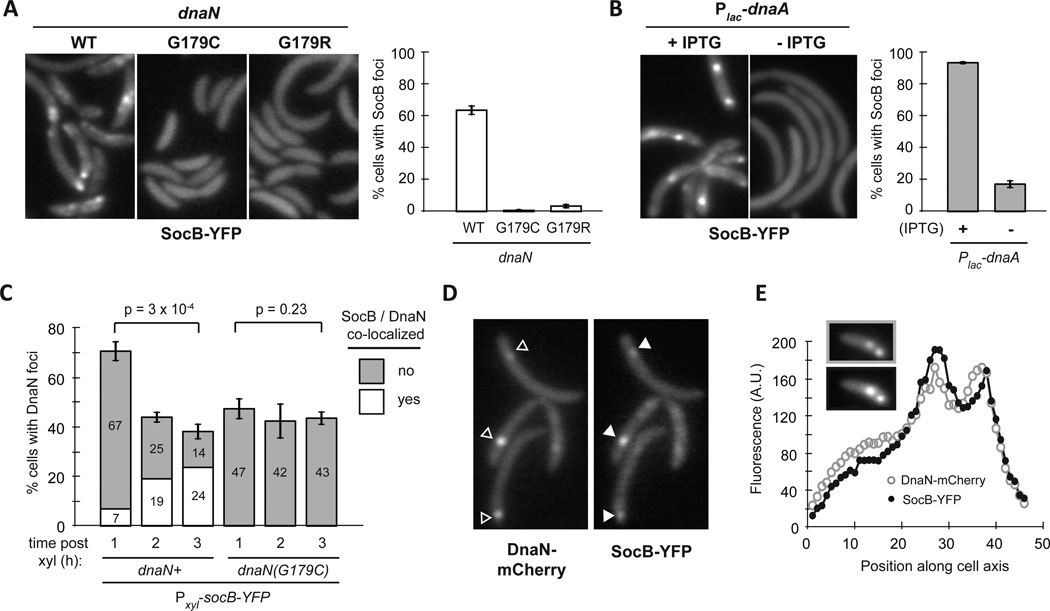
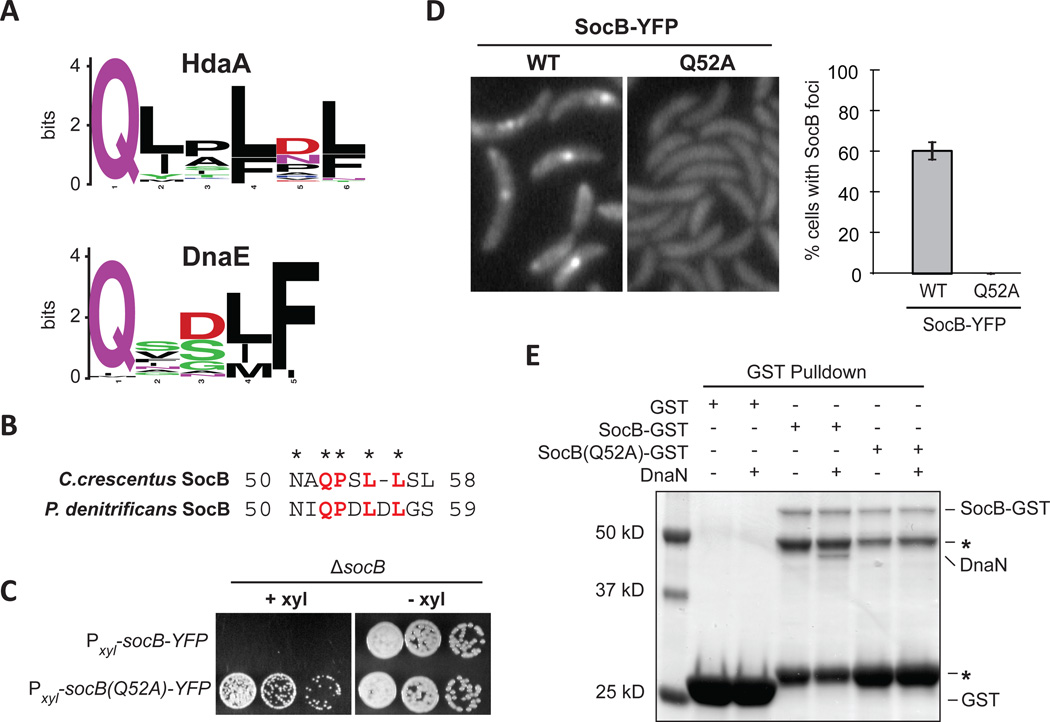
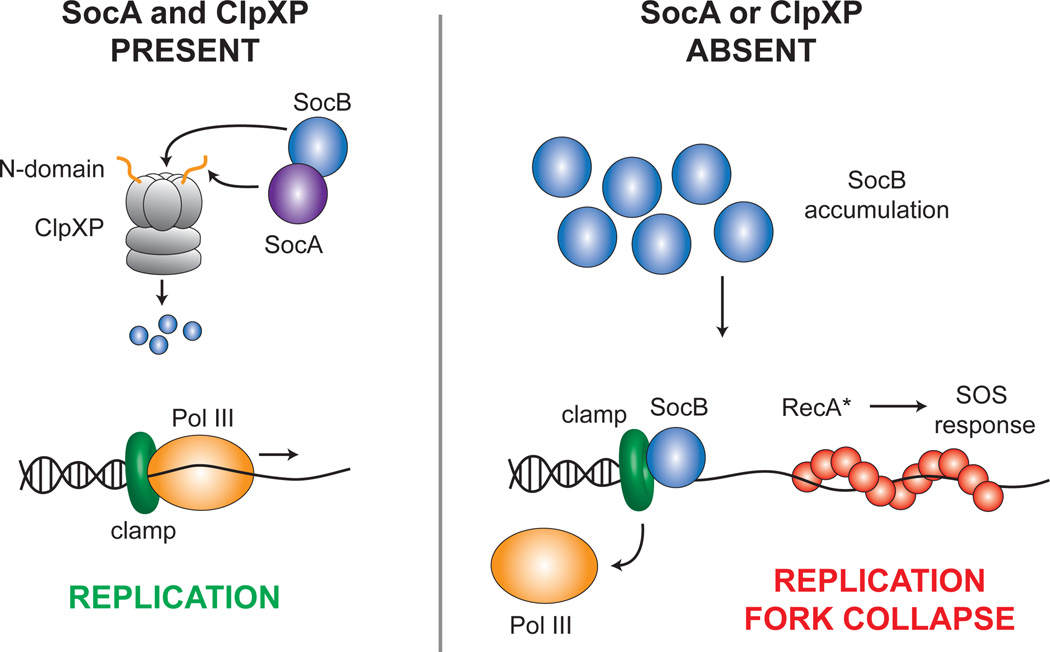
Toxin-antitoxin (TA) systems are ubiquitous on bacterial chromosomes, yet the mechanisms regulating their activity and the molecular targets of toxins remain incompletely defined. Here, we identify SocAB, an atypical TA system in Caulobacter crescentus. Unlike canonical TA systems, the toxin SocB is unstable and constitutively degraded by the protease ClpXP; this degradation requires the antitoxin, SocA, as a proteolytic adaptor. We find that the toxin, SocB, blocks replication elongation through an interaction with the sliding clamp, driving replication fork collapse. Mutations that suppress SocB toxicity map to either the hydrophobic cleft on the clamp that binds DNA polymerase III or a clamp-binding motif in SocB. Our findings suggest that SocB disrupts replication by outcompeting other clamp-binding proteins. Collectively, our results expand the diversity of mechanisms employed by TA systems to regulate toxin activity and inhibit bacterial growth, and they suggest that inhibiting clamp function may be a generalizable antibacterial strategy.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
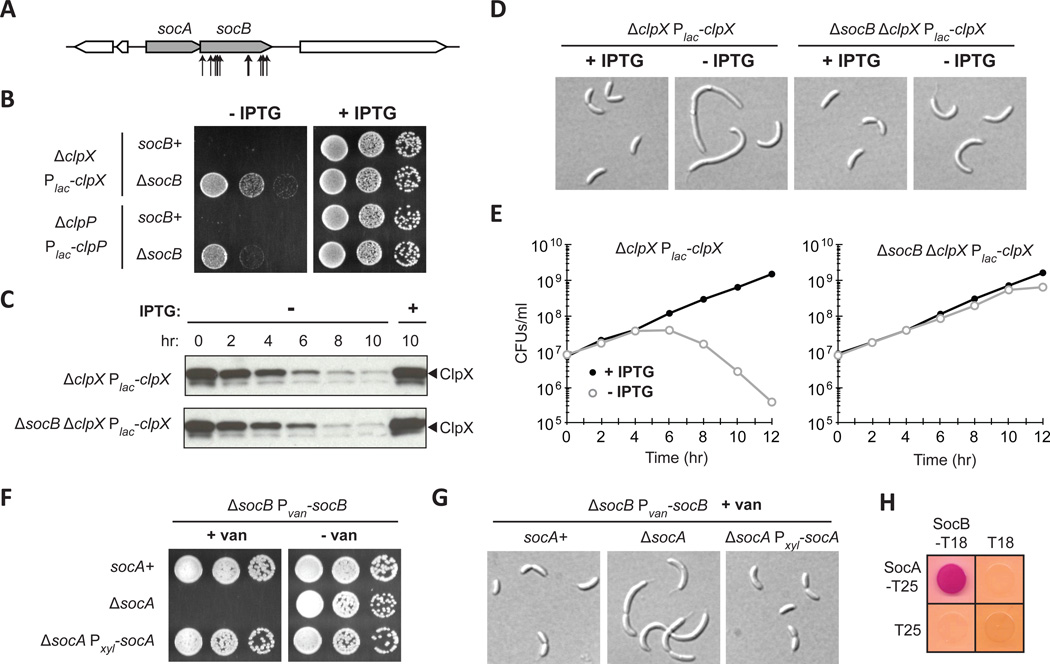
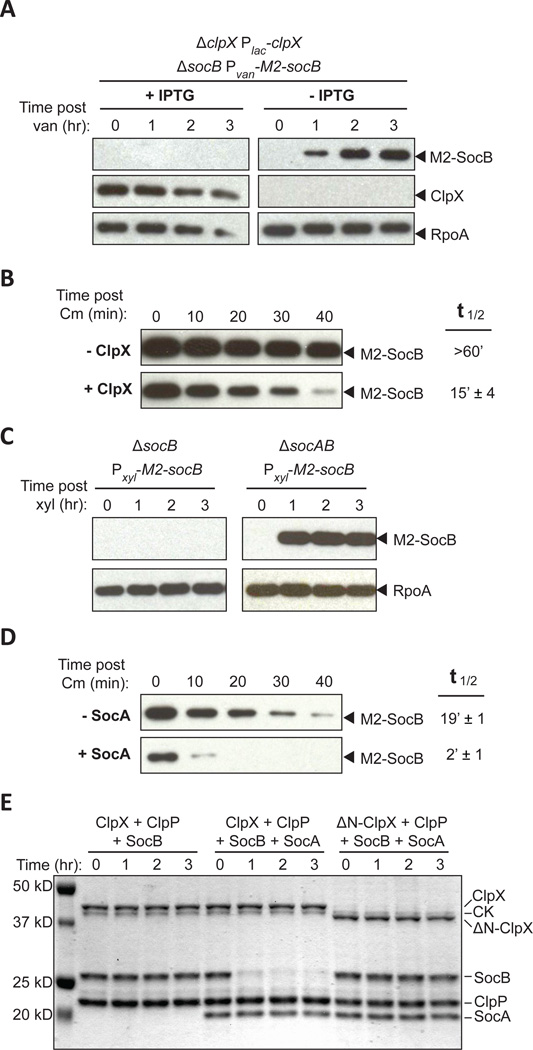
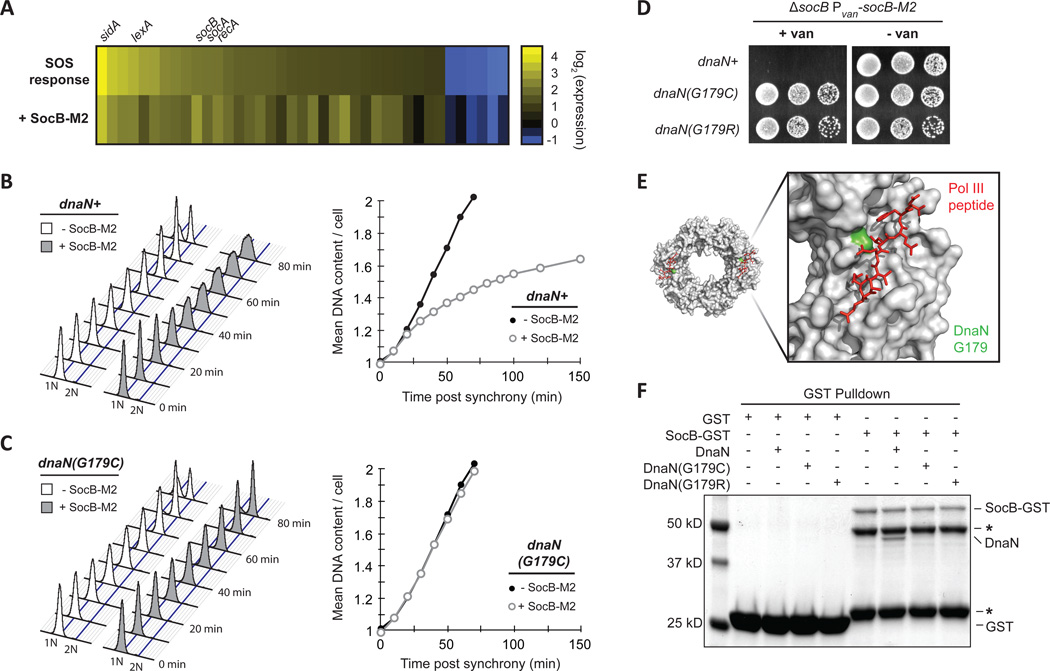
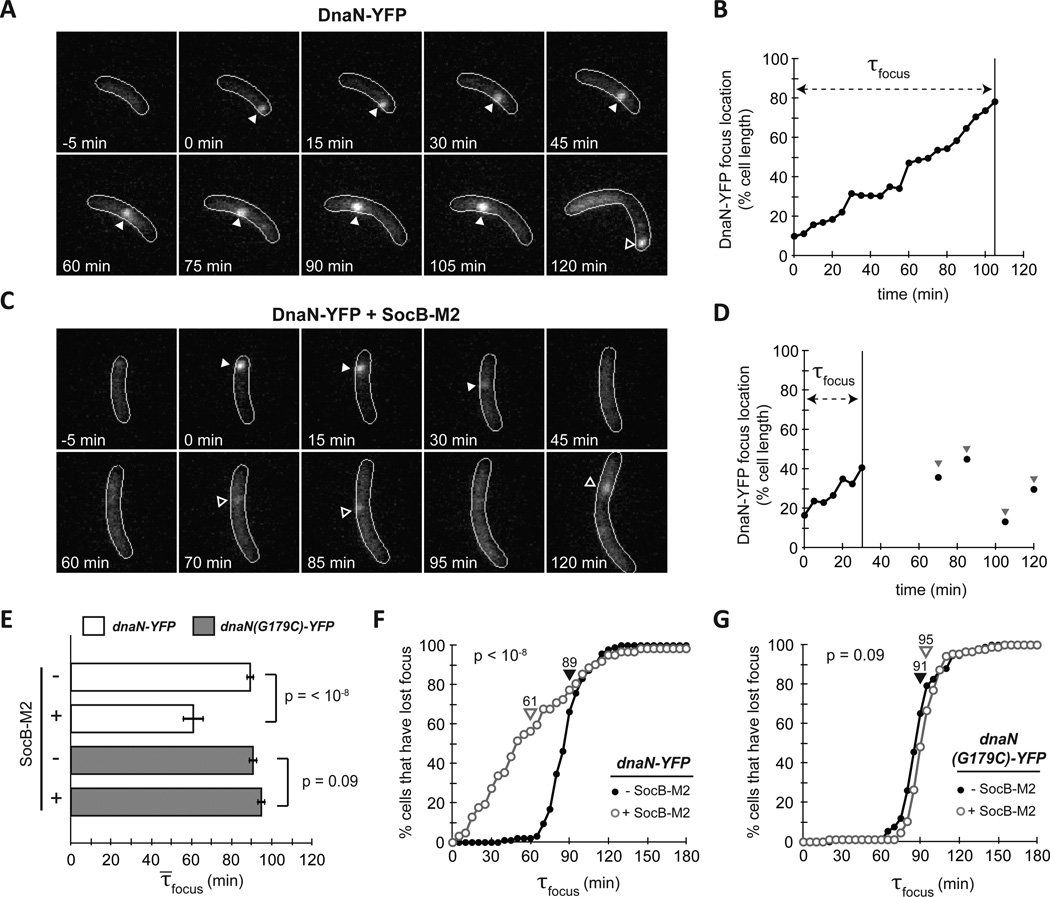
Comment in
-
Preventing bacterial suicide: a novel toxin-antitoxin strategy.Mol Cell. 2013 Dec 12;52(5):611-2. doi: 10.1016/j.molcel.2013.11.018. Mol Cell. 2013. PMID: 24332174 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
