

The PAR polarity complex and cerebellar granule neuron migration
- PMID: 24243103
- PMCID: PMC8202091
- DOI: 10.1007/978-94-007-7687-6_7
The PAR polarity complex and cerebellar granule neuron migration
Abstract
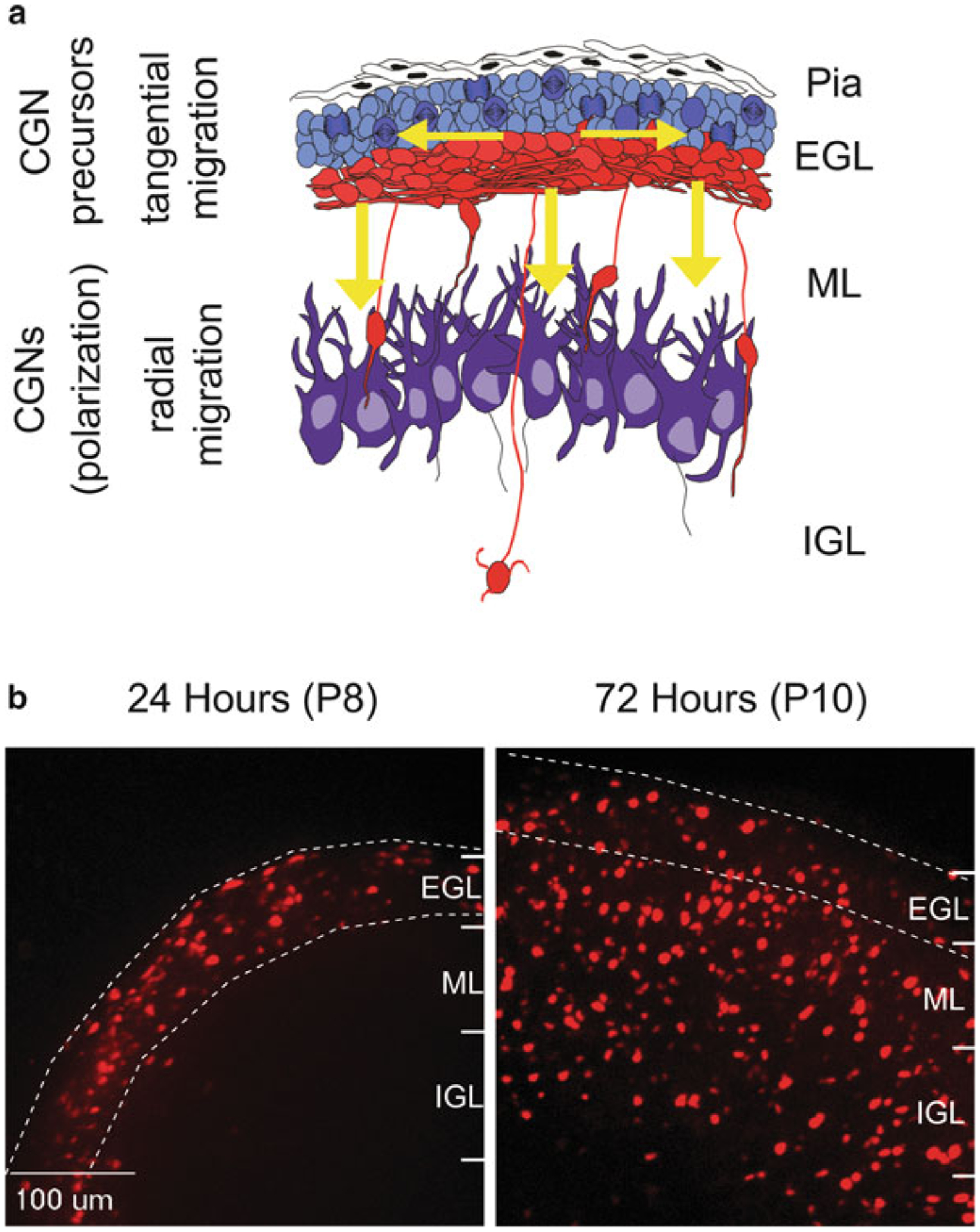
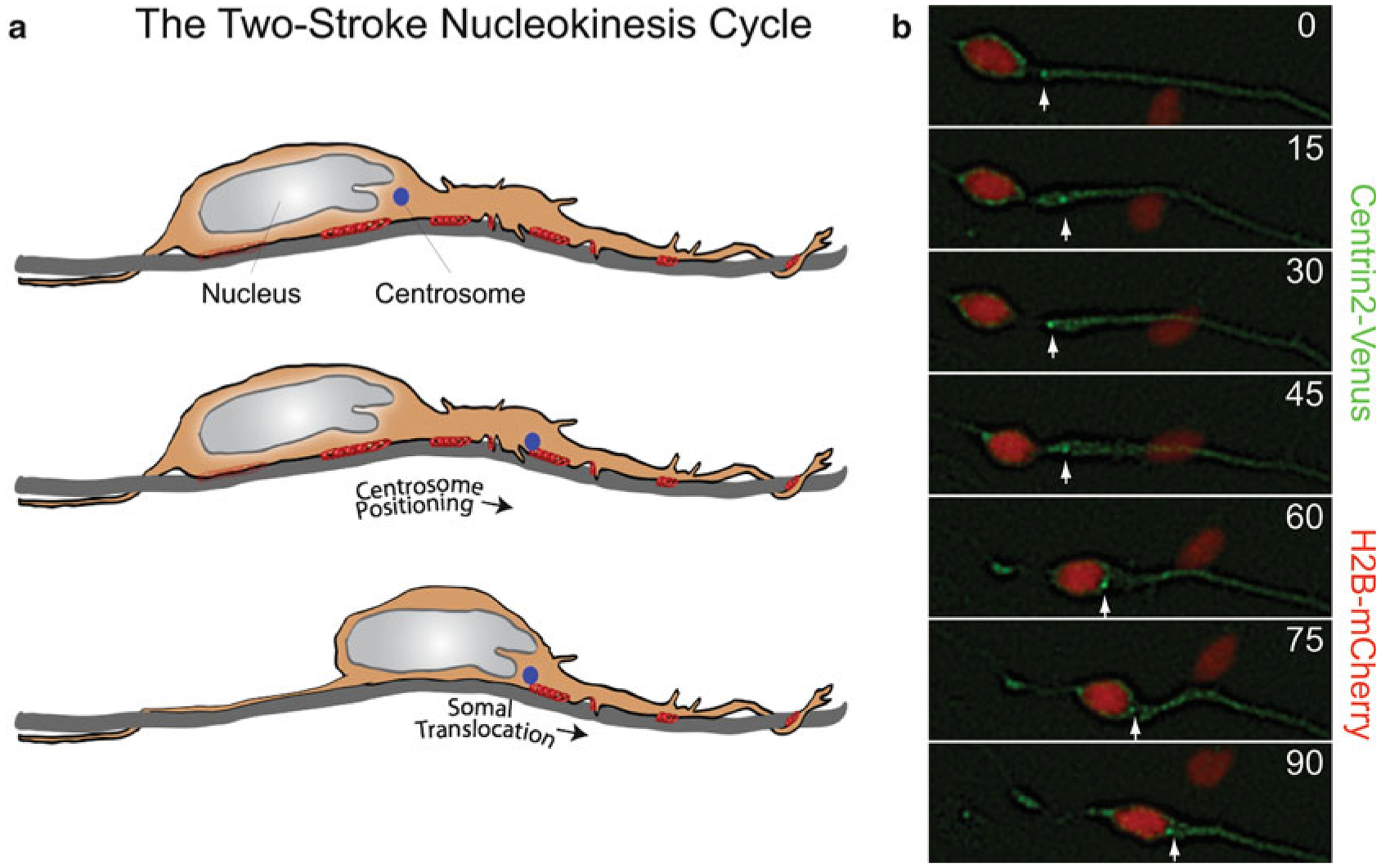
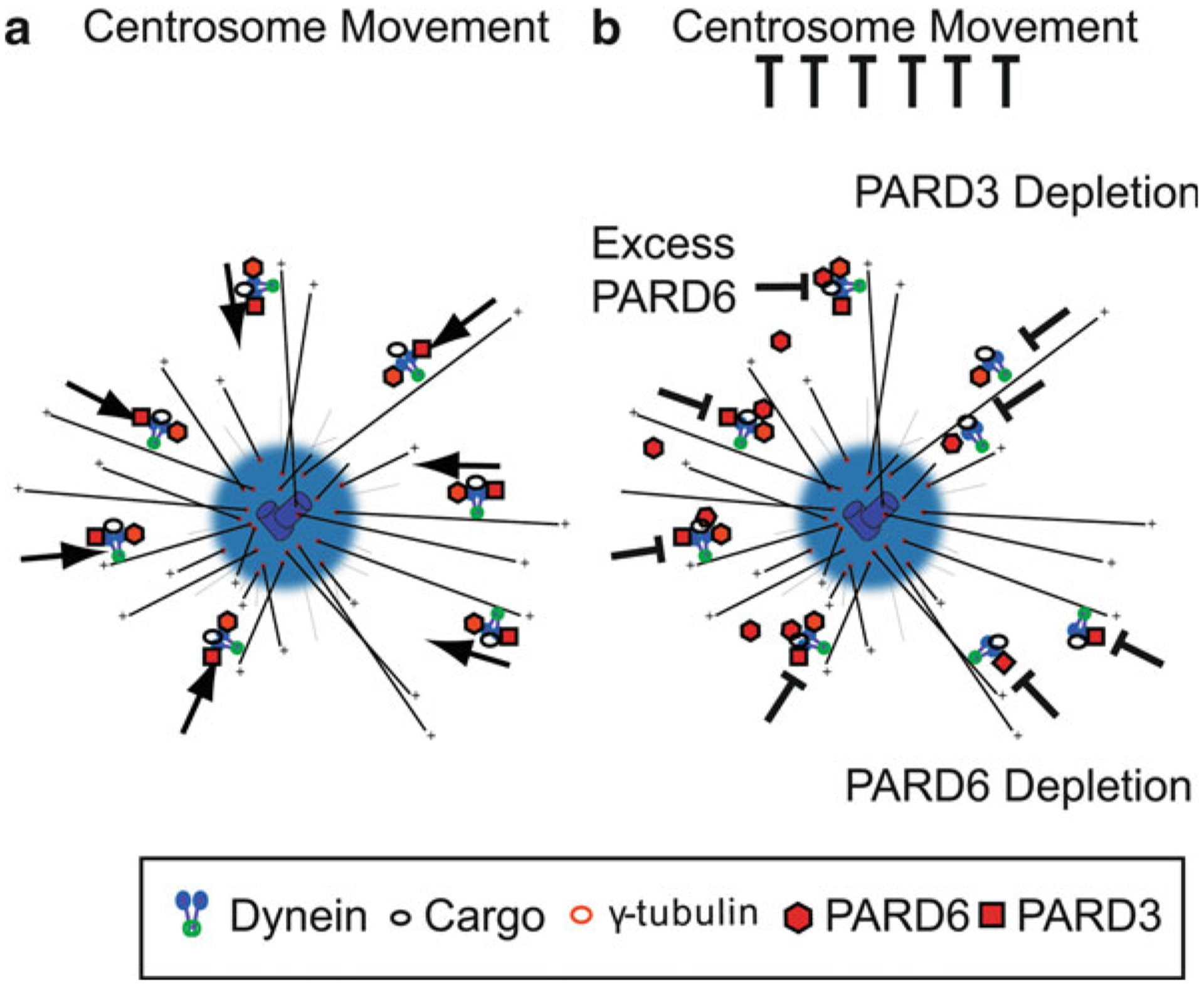
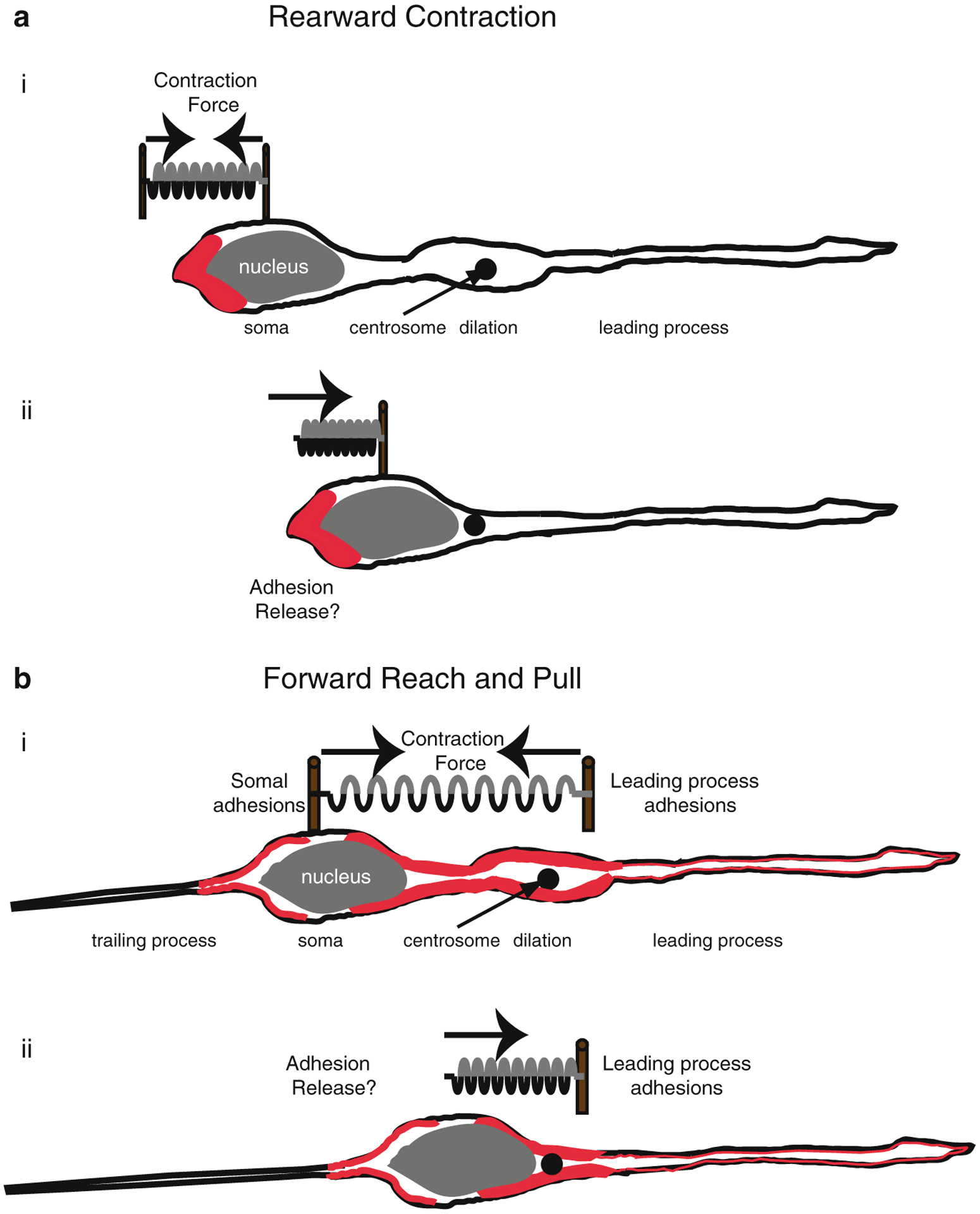
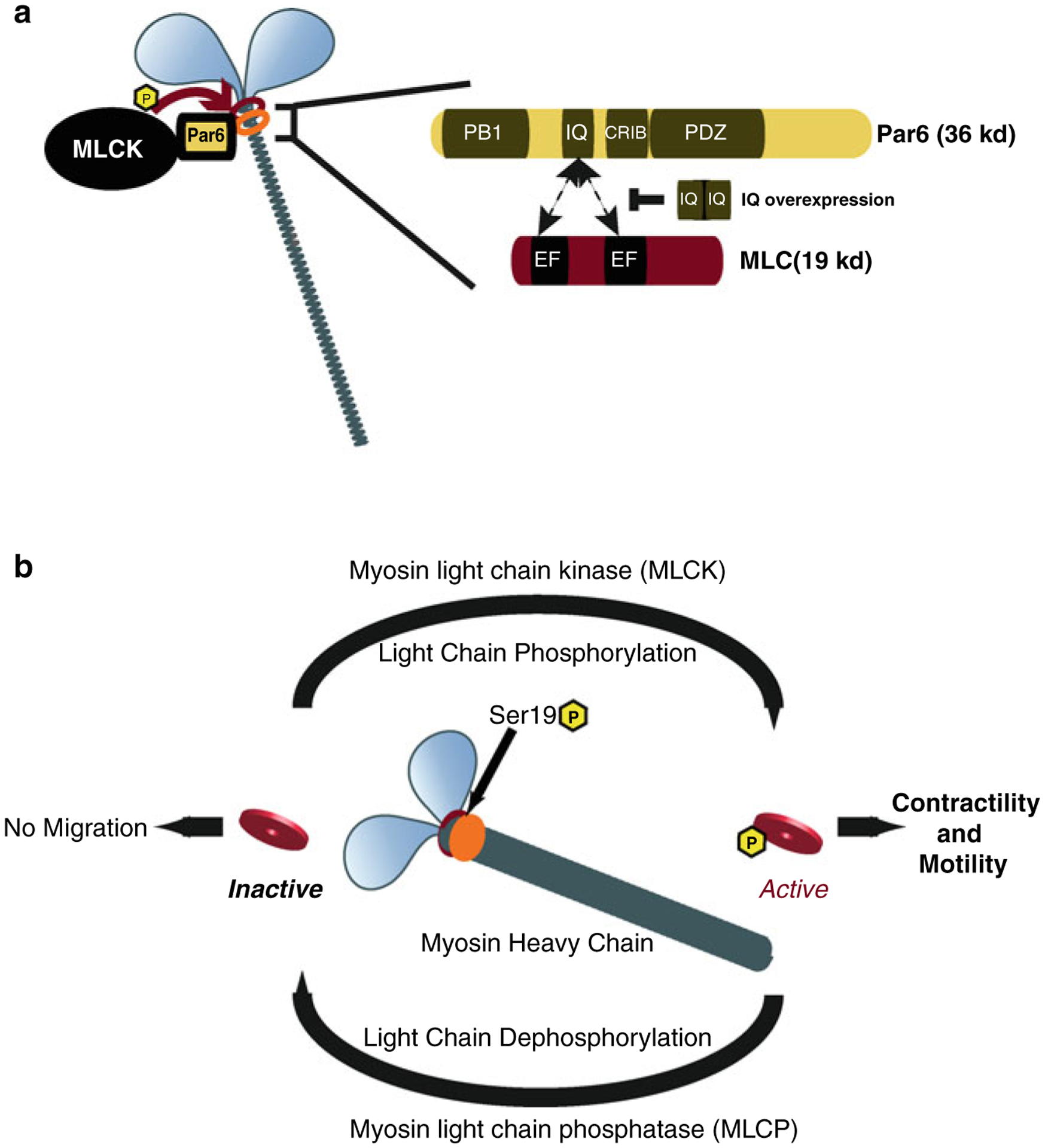
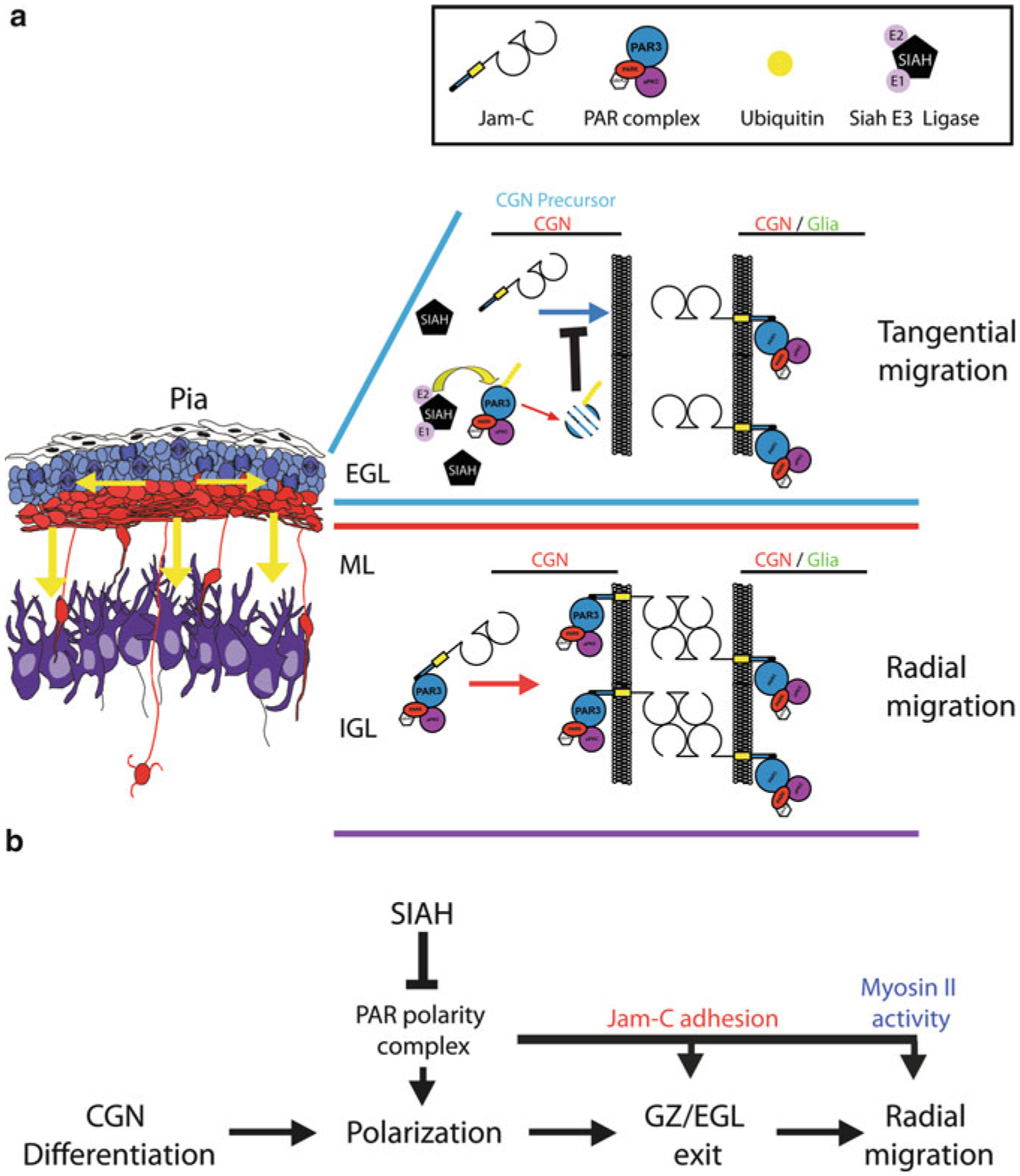
Proper migration of neurons is one of the most important aspects of early brain development. After neuronal progenitors are born in their respective germinal niches, they must migrate to their final locations to form precise neural circuits. A majority of migrating neurons move by associating and disassociating with glial fibers, which serve as scaffolding for the developing brain. Cerebellar granule neurons provide a model system for examination of the mechanisms of neuronal migration in dissociated and slice culture systems; the ability to purify these cells allows migration assays to be paired with genetic, molecular, and biochemical findings. CGNs migrate in a highly polarized fashion along radial glial fibers, using a two-stroke nucleokinesis cycle. The PAR polarity complex of PARD3, PARD6, and an atypical protein kinase C (aPKC) regulate several aspects of neuronal migration. The PAR polarity complex regulates the coordinated movements of the centrosome and soma during nucleokinesis, and also the stability of the microtubule cytoskeleton during migration. PAR proteins coordinate actomyosin dynamics in the leading process of migrating neurons, which are required for migration. The PAR complex also controls the cell-cell adhesions made by migrating neurons along glial cells, and through this mechanism regulates germinal zone exit during prenatal brain development. These findings suggest that the PAR complex coordinates the movement of multiple cellular elements as neurons migrate and that further examination of PAR complex effectors will not only provide novel insights to address fundamental challenges to the field but also expand our understanding of how the PAR complex functions at the molecular level.
Figures
References
-
- Allen KM, Gleeson JG, Shoup SM, Walsh CA (1998) A YAC contig in Xq22. 3-q23, from DXS287 to DXS8088, spanning the brain-specific genes doublecortin (DCX) and PAK3. Genomics 52(2):214–218 - PubMed
-
- Bellion A, Baudoin JP, Alvarez C, Bornens M, Metin C (2005) Nucleokinesis in tangentially migrating neurons comprises two alternating phases: forward migration of the Golgi/centrosome associated with centrosome splitting and myosin contraction at the rear. J Neurosci 25(24):5691–5699. doi:25/24/5691 [pii] 10.1523/JNEUROSCI.1030-05.2005 - DOI - PMC - PubMed
-
- Bertet C, Sulak L, Lecuit T (2004) Myosin-dependent junction remodelling controls planar cell intercalation and axis elongation. Nature 429(6992):667–671. doi: http://www.nature.com/nature/journal/v429/n6992/suppinfo/nature02590_S1.... - PubMed
-
- Chen S, Chen J, Shi H, Wei M, Castaneda-Castellanos David R, Bultje Ronald S, Pei X, Kriegstein Arnold R, Zhang M, Shi S-H (2013) Regulation of microtubule stability and organization by mammalian Par3 in specifying neuronal polarity. Dev Cell 24(1):26–40. doi: 10.1016/j.devcel.2012.11.014 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
