

Molecular characterization of a human matrix attachment region epigenetic regulator
- PMID: 24244463
- PMCID: PMC3828356
- DOI: 10.1371/journal.pone.0079262
Molecular characterization of a human matrix attachment region epigenetic regulator
Abstract
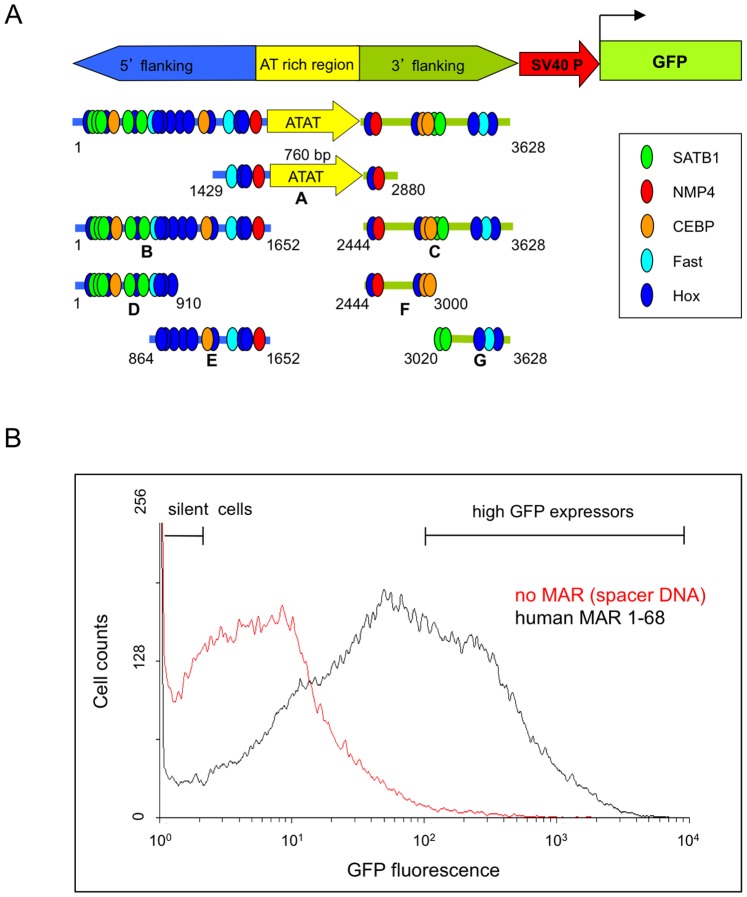
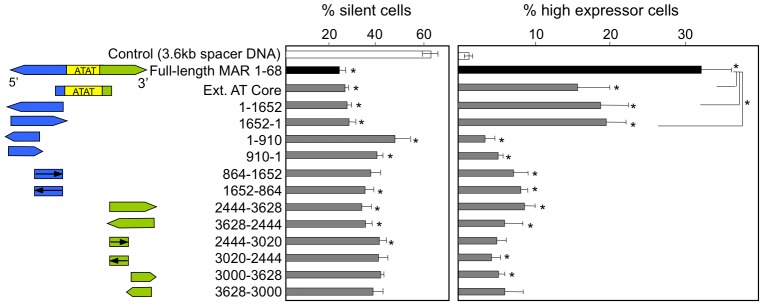
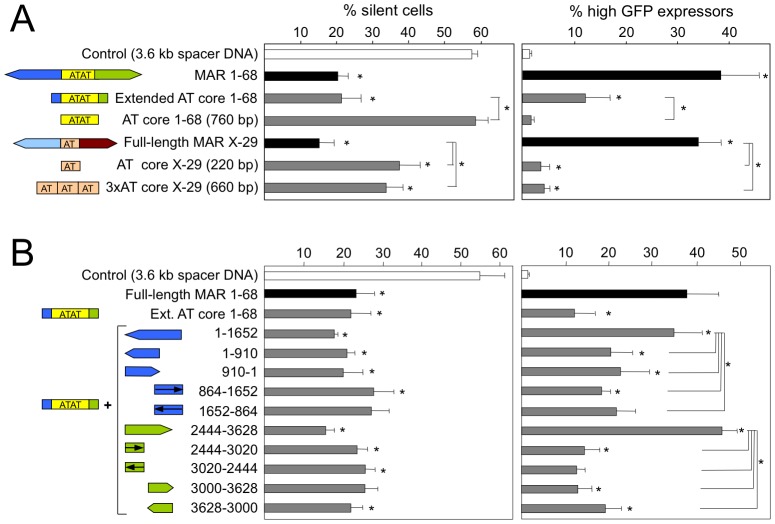
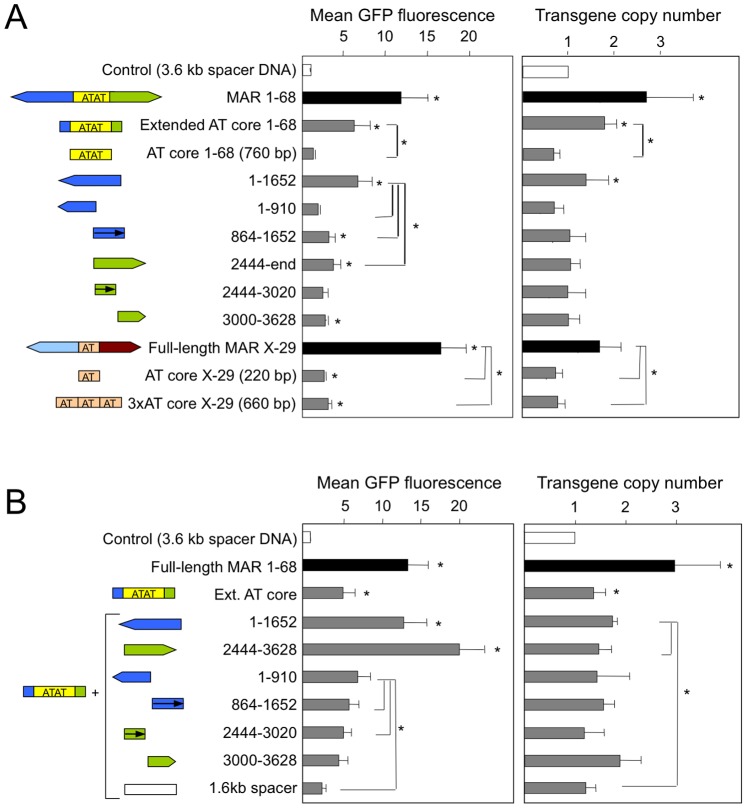
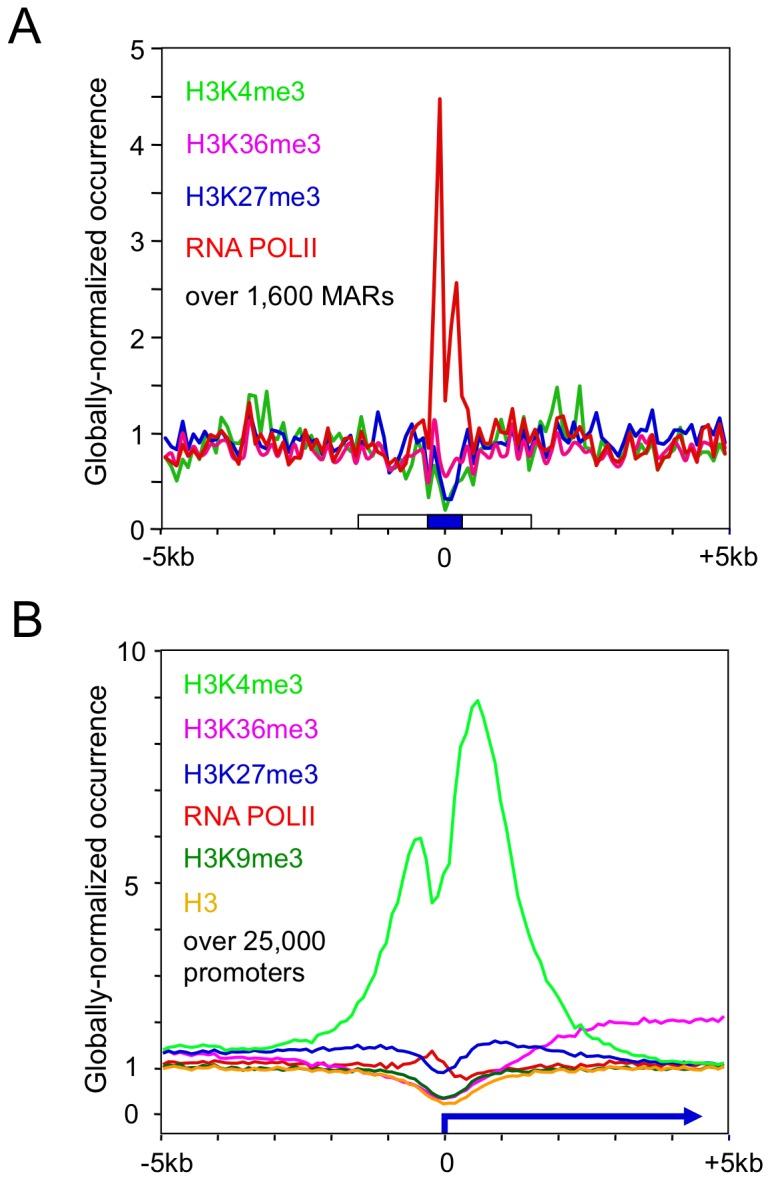
Matrix attachment regions (MAR) generally act as epigenetic regulatory sequences that increase gene expression, and they were proposed to partition chromosomes into loop-forming domains. However, their molecular mode of action remains poorly understood. Here, we assessed the possible contribution of the AT-rich core and adjacent transcription factor binding motifs to the transcription augmenting and anti-silencing effects of human MAR 1-68. Either flanking sequences together with the AT-rich core were required to obtain the full MAR effects. Shortened MAR derivatives retaining full MAR activity were constructed from combinations of the AT-rich sequence and multimerized transcription factor binding motifs, implying that both transcription factors and the AT-rich microsatellite sequence are required to mediate the MAR effect. Genomic analysis indicated that MAR AT-rich cores may be depleted of histones and enriched in RNA polymerase II, providing a molecular interpretation of their chromatin domain insulator and transcriptional augmentation activities.
Conflict of interest statement
Figures
References
-
- Heng HH, Goetze S, Ye CJ, Liu G, Stevens JB, et al. (2004) Chromatin loops are selectively anchored using scaffold/matrix-attachment regions. J Cell Sci 117: 999–1008. - PubMed
-
- Felsenfeld G, Burgess-Beusse B, Farrell C, Gaszner M, Ghirlando R, et al. (2004) Chromatin boundaries and chromatin domains. Cold Spring Harb Symp Quant Biol 69: 245–250. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
