

MuSK myasthenia gravis IgG4 disrupts the interaction of LRP4 with MuSK but both IgG4 and IgG1-3 can disperse preformed agrin-independent AChR clusters
- PMID: 24244707
- PMCID: PMC3820634
- DOI: 10.1371/journal.pone.0080695
MuSK myasthenia gravis IgG4 disrupts the interaction of LRP4 with MuSK but both IgG4 and IgG1-3 can disperse preformed agrin-independent AChR clusters
Abstract
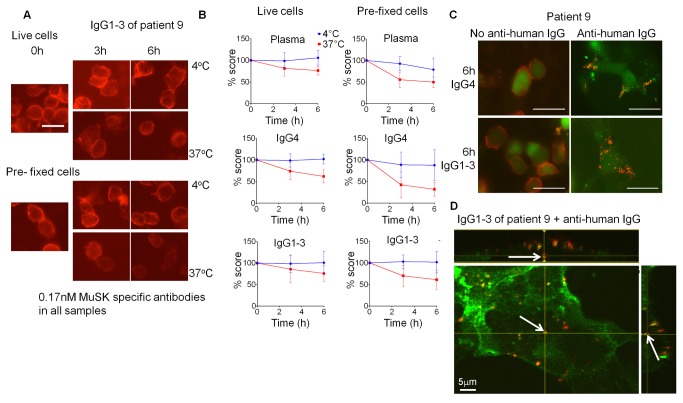
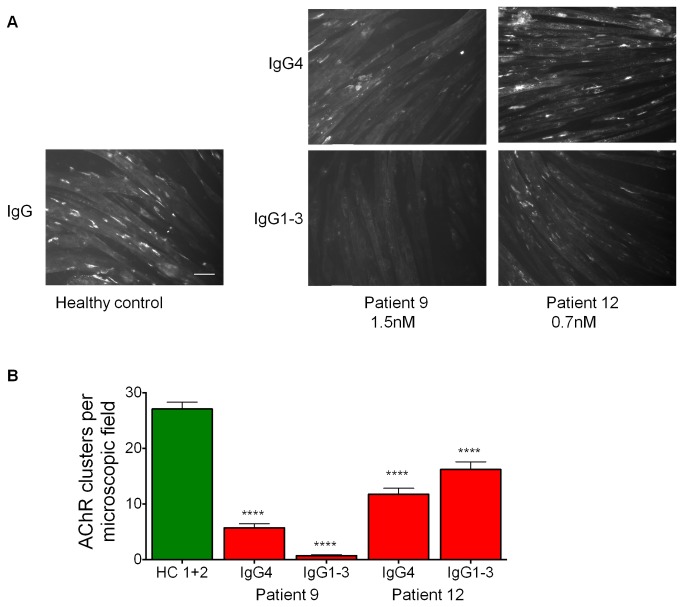
A variable proportion of patients with generalized myasthenia gravis (MG) have autoantibodies to muscle specific tyrosine kinase (MuSK). During development agrin, released from the motor nerve, interacts with low density lipoprotein receptor-related protein-4 (LRP4), which then binds to MuSK; MuSK interaction with the intracellular protein Dok7 results in clustering of the acetylcholine receptors (AChRs) on the postsynaptic membrane. In mature muscle, MuSK helps maintain the high density of AChRs at the neuromuscular junction. MuSK antibodies are mainly IgG4 subclass, which does not activate complement and can be monovalent, thus it is not clear how the antibodies cause disruption of AChR numbers or function to cause MG. We hypothesised that MuSK antibodies either reduce surface MuSK expression and/or inhibit the interaction with LRP4. We prepared MuSK IgG, monovalent Fab fragments, IgG1-3 and IgG4 fractions from MuSK-MG plasmas. We asked whether the antibodies caused endocytosis of MuSK in MuSK-transfected cells or if they inhibited binding of LRP4 to MuSK in co-immunoprecipitation experiments. In parallel, we investigated their ability to reduce AChR clusters in C2C12 myotubes induced by a) agrin, reflecting neuromuscular development, and b) by Dok7- overexpression, producing AChR clusters that more closely resemble the adult neuromuscular synapse. Total IgG, IgG4 or IgG1-3 MuSK antibodies were not endocytosed unless cross-linked by divalent anti-human IgG. MuSK IgG, Fab fragments and IgG4 inhibited the binding of LRP4 to MuSK and reduced agrin-induced AChR clustering in C2C12 cells. By contrast, IgG1-3 antibodies did not inhibit LRP4-MuSK binding but, surprisingly, did inhibit agrin-induced clustering. Moreover, both IgG4 and IgG1-3 preparations dispersed agrin-independent AChR clusters in Dok7-overexpressing C2C12 cells. Thus interference by IgG4 antibodies of the LRP4-MuSK interaction will be one pathogenic mechanism of MuSK antibodies, but IgG1-3 MuSK antibodies will also contribute to the reduced AChR density and neuromuscular dysfunction in myasthenia patients with MuSK antibodies.
Conflict of interest statement
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
