

Bacterial natural transformation by highly fragmented and damaged DNA
- PMID: 24248361
- PMCID: PMC3856829
- DOI: 10.1073/pnas.1315278110
Bacterial natural transformation by highly fragmented and damaged DNA
Abstract
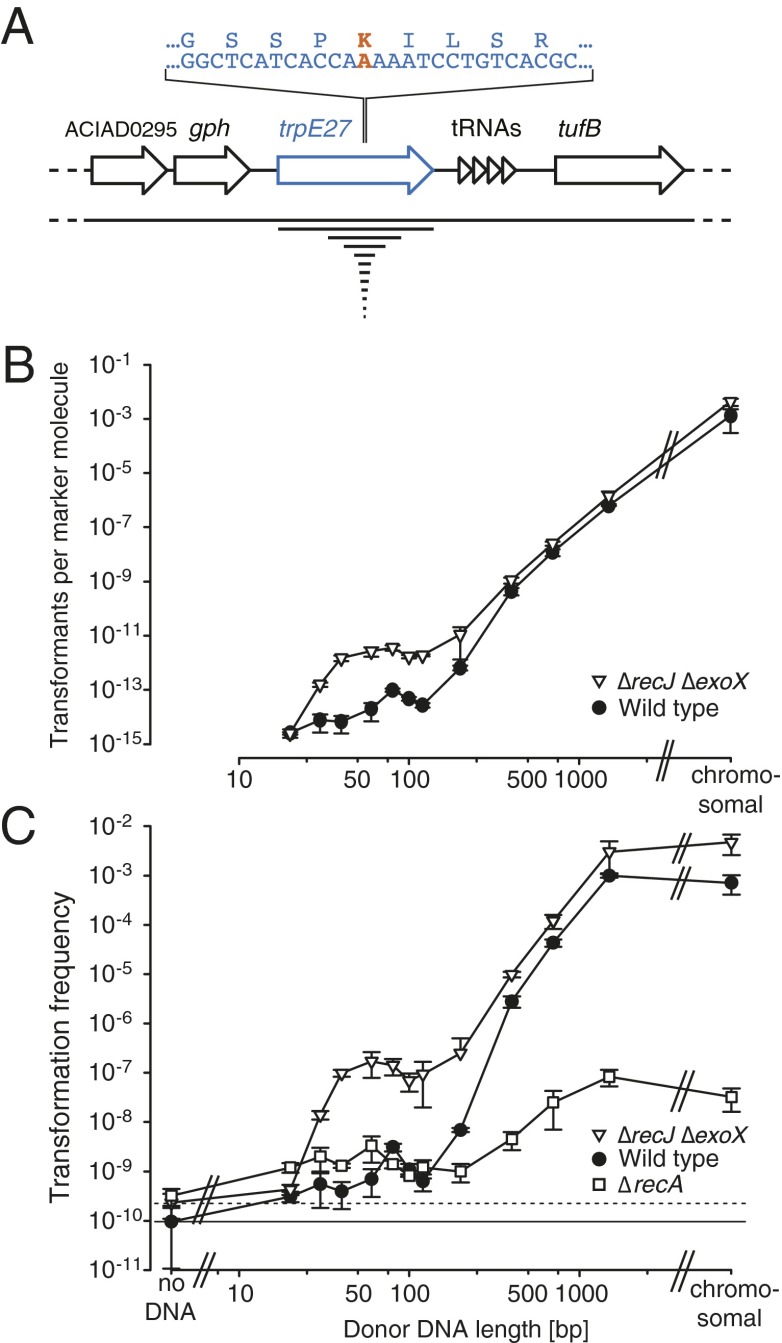
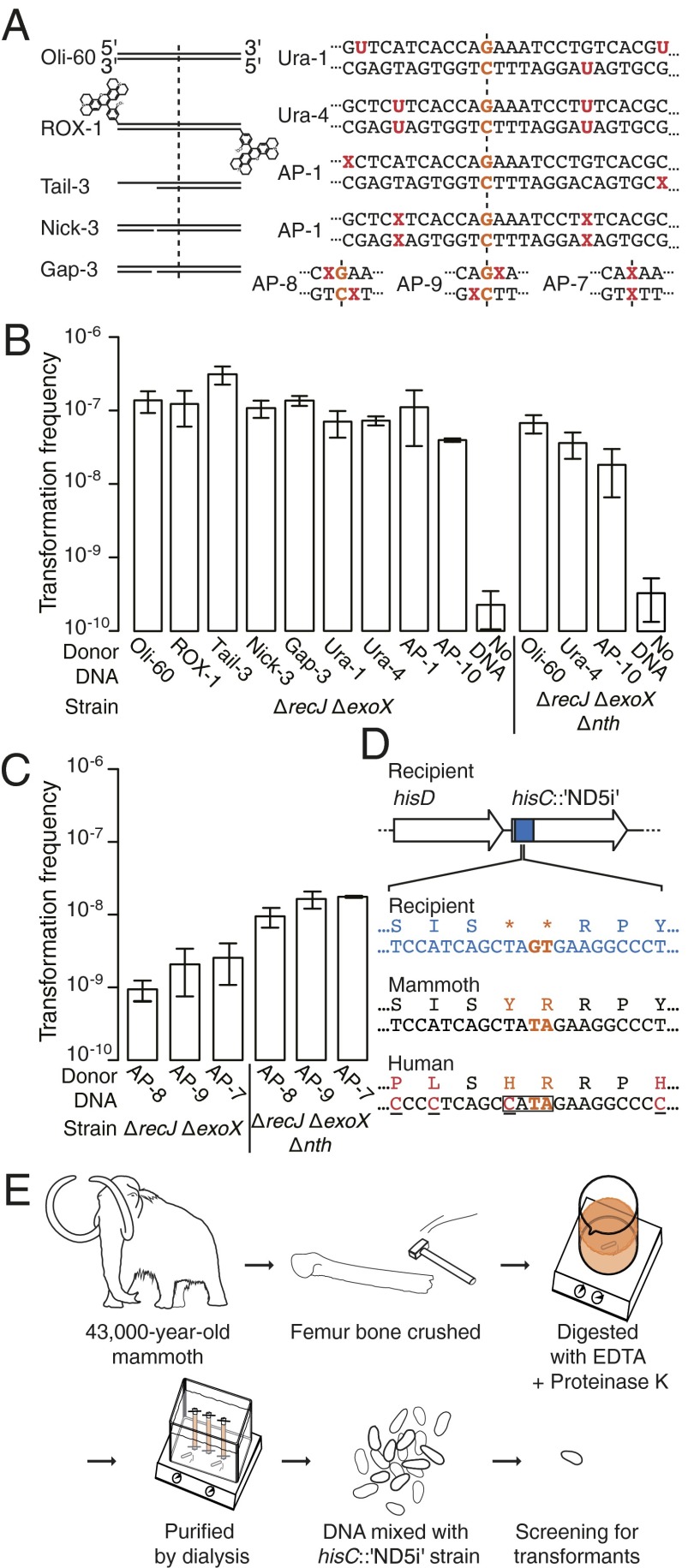
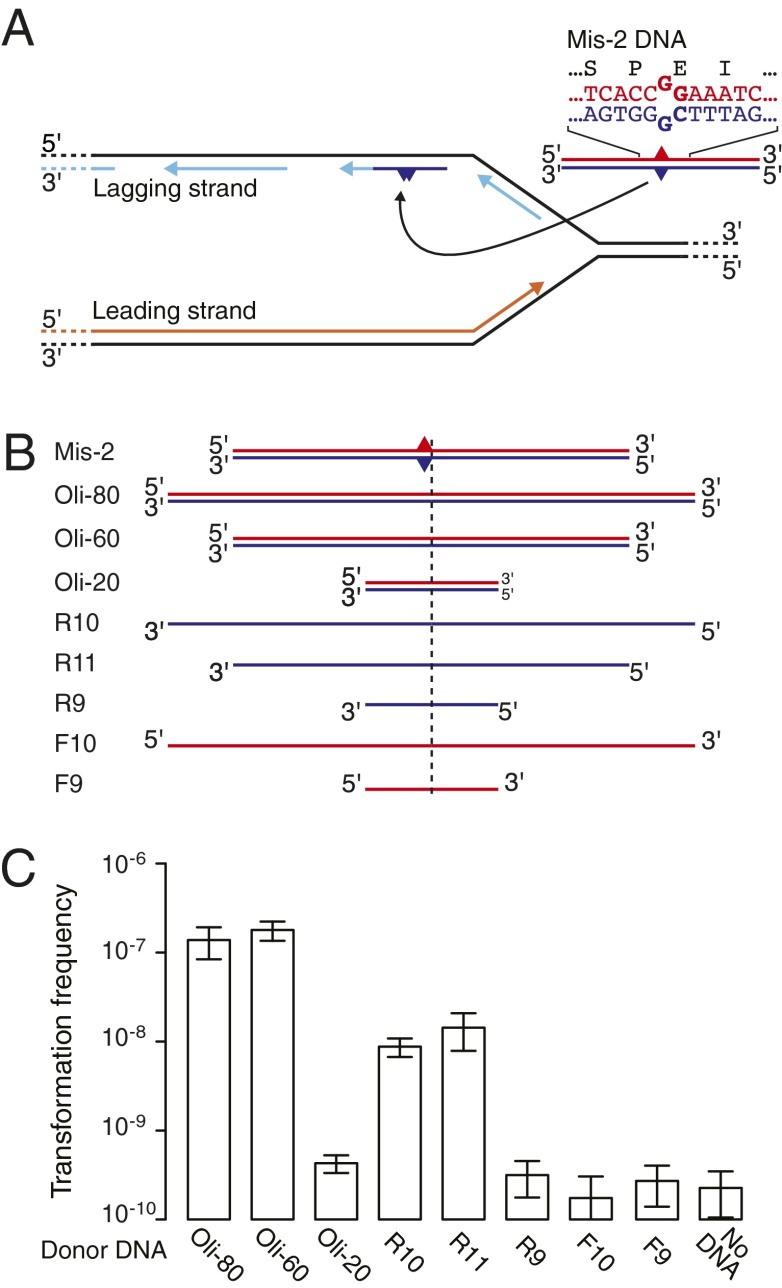
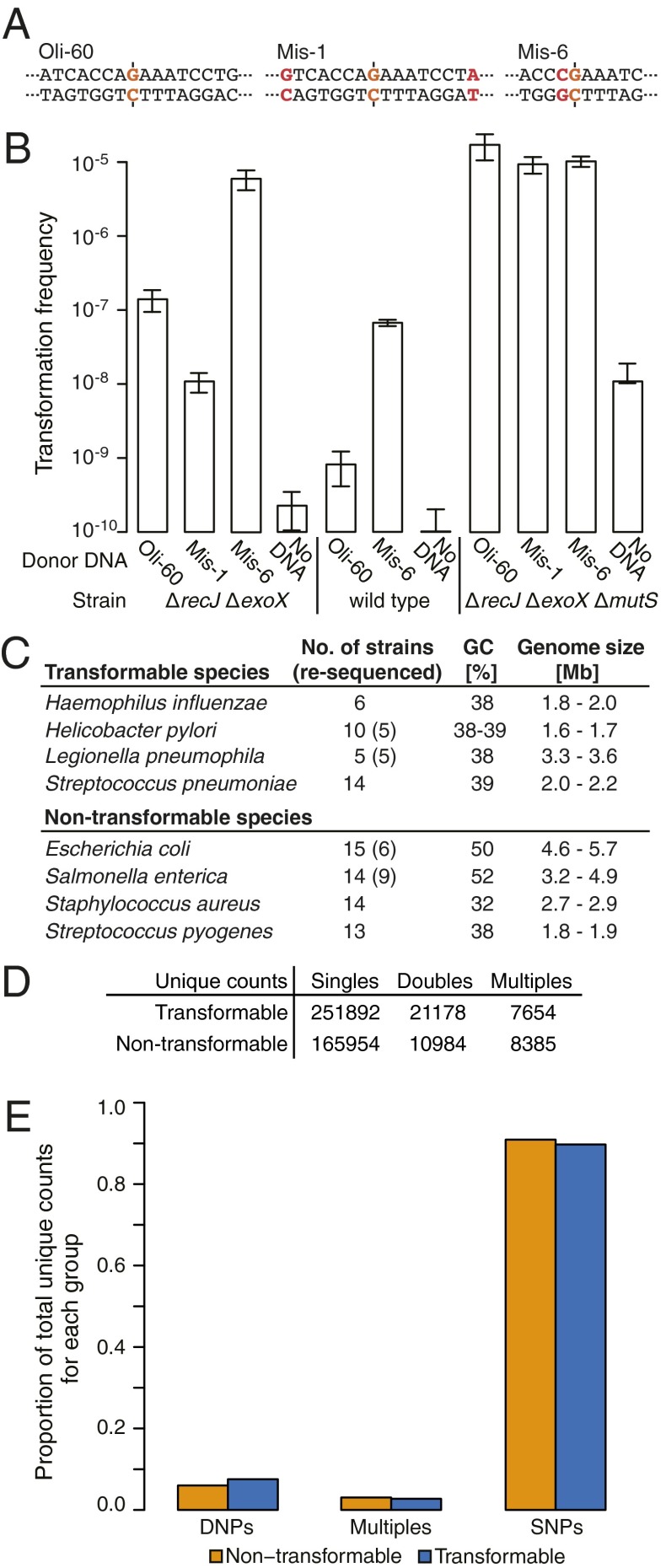
DNA molecules are continuously released through decomposition of organic matter and are ubiquitous in most environments. Such DNA becomes fragmented and damaged (often <100 bp) and may persist in the environment for more than half a million years. Fragmented DNA is recognized as nutrient source for microbes, but not as potential substrate for bacterial evolution. Here, we show that fragmented DNA molecules (≥ 20 bp) that additionally may contain abasic sites, cross-links, or miscoding lesions are acquired by the environmental bacterium Acinetobacter baylyi through natural transformation. With uptake of DNA from a 43,000-y-old woolly mammoth bone, we further demonstrate that such natural transformation events include ancient DNA molecules. We find that the DNA recombination is RecA recombinase independent and is directly linked to DNA replication. We show that the adjacent nucleotide variations generated by uptake of short DNA fragments escape mismatch repair. Moreover, double-nucleotide polymorphisms appear more common among genomes of transformable than nontransformable bacteria. Our findings reveal that short and damaged, including truly ancient, DNA molecules, which are present in large quantities in the environment, can be acquired by bacteria through natural transformation. Our findings open for the possibility that natural genetic exchange can occur with DNA up to several hundreds of thousands years old.
Keywords: DNA degradation; anachronistic evolution; early life; horizontal gene transfer; microbial evolution.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Nielsen KM, Johnsen PJ, Bensasson D, Daffonchio D. Release and persistence of extracellular DNA in the environment. Environ Biosafety Res. 2007;6(1-2):37–53. - PubMed
-
- Lindahl T. Instability and decay of the primary structure of DNA. Nature. 1993;362(6422):709–715. - PubMed
-
- Pietramellara G, Ascher J, Borgogni F, Ceccherini GGMT, Nannipieri P. Extracellular DNA in soil and sediment: Fate and ecological relevance. Biol Fertil Soils. 2009;45(3):219–235.
-
- Willerslev E, et al. Long-term persistence of bacterial DNA. Curr Biol. 2004;14(1):R9–R10. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
