

Enzymatic production of mono-ubiquitinated proteins for structural studies: The example of the Josephin domain of ataxin-3
- PMID: 24251111
- PMCID: PMC3829987
- DOI: 10.1016/j.fob.2013.10.005
Enzymatic production of mono-ubiquitinated proteins for structural studies: The example of the Josephin domain of ataxin-3
Abstract
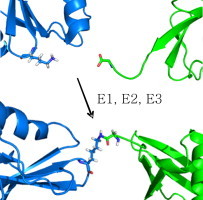
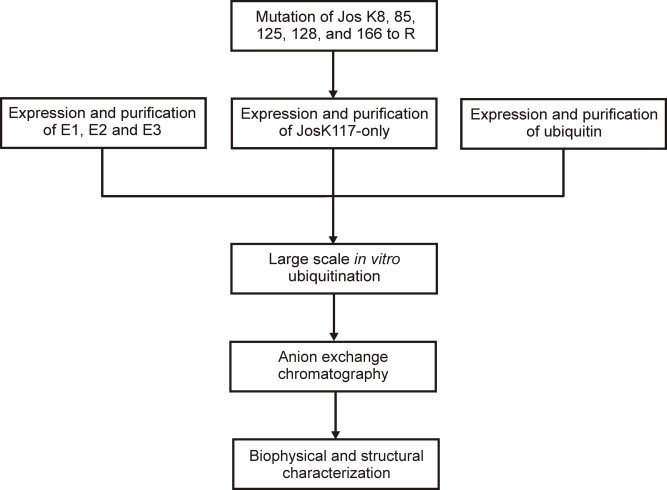
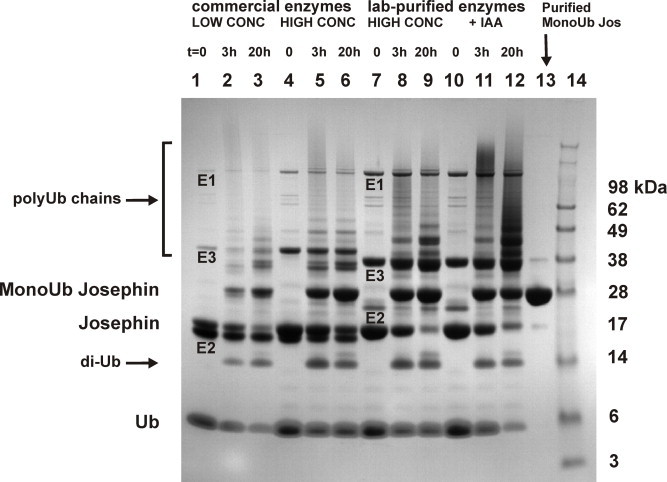
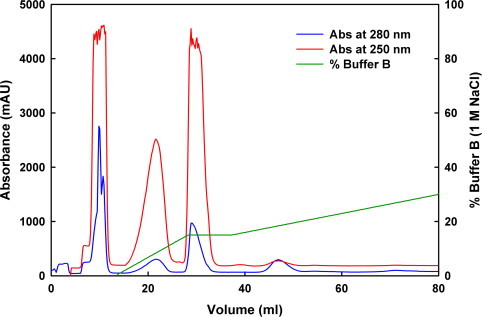
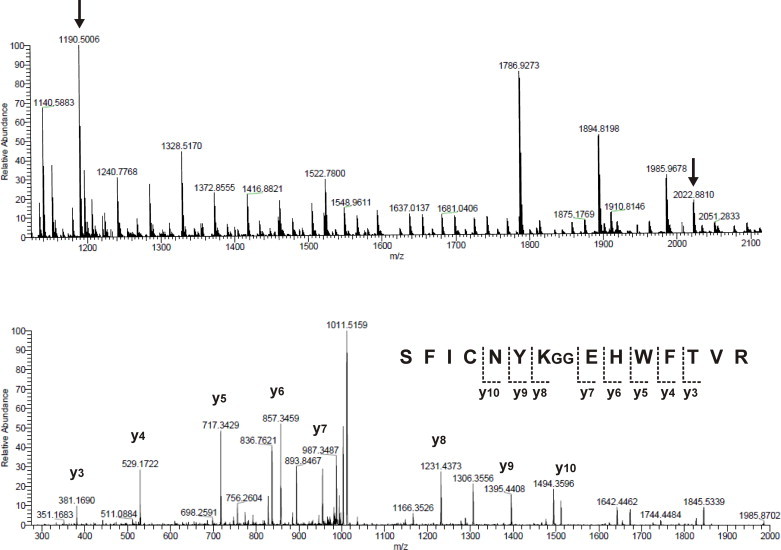
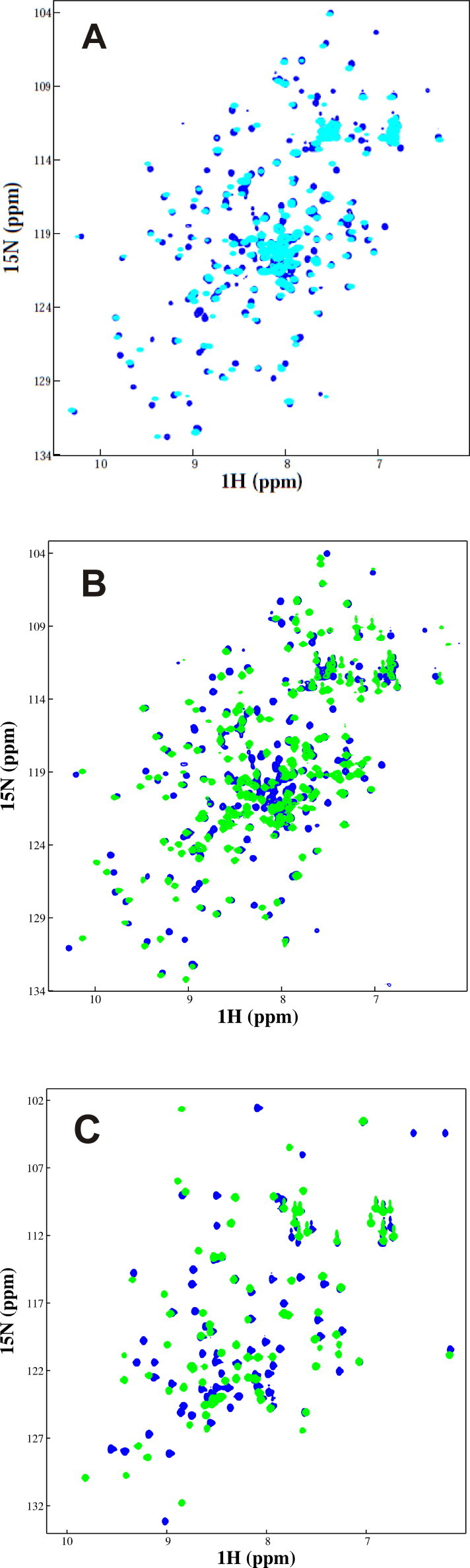
Protein ubiquitination occurs through formation of an isopeptide bond between the C-terminal glycine of ubiquitin (Ub) and the ɛ-amino group of a substrate lysine residue. This post-translational modification, which occurs through the attachment of single and/or multiple copies of mono-ubiquitin and poly-ubiquitin chains, is involved in crucial cellular events such as protein degradation, cell-cycle regulation and DNA repair. The abnormal functioning of ubiquitin pathways is also implicated in the pathogenesis of several human diseases ranging from cancer to neurodegeneration. However, despite the undoubted biological importance, understanding the molecular basis of how ubiquitination regulates different pathways has up to now been strongly limited by the difficulty of producing the amounts of highly homogeneous samples that are needed for a structural characterization by X-ray crystallography and/or NMR. Here, we report on the production of milligrams of highly pure Josephin mono-ubiquitinated on lysine 117 through large scale in vitro enzymatic ubiquitination. Josephin is the catalytic domain of ataxin-3, a protein responsible for spinocerebellar ataxia type 3. Ataxin-3 is the first deubiquitinating enzyme (DUB) reported to be activated by mono-ubiquitination. We demonstrate that the samples produced with the described method are correctly folded and suitable for structural studies. The protocol allows facile selective labelling of the components. Our results provide an important proof-of-concept that may pave the way to new approaches to the in vitro study of ubiquitinated proteins.
Keywords: ATP, adenosine triphosphate; DTT, dithiothreitol; DUB, deubiquitinating enzyme; Deubiquitinating enzyme; GST, glutathione-S-transferase; HSQC, heteronuclear single quantum coherence; IAA, iodoacetamide; Isopeptide bond; JosK117-only, Josephin mutant in which all lysines but K117 are mutated; Josephin; MS/MS tandem, mass spectrometry; Machado–Joseph disease; NMR, nuclear magnetic resonance; PDB, Protein Data Bank; Post-translational modification; SDS–PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis; Spinocerebellar ataxia type 3; Tris–HCl, 2-amino-2-(hydroxymethyl)-1,3-propanediol hydrochloride; Ubiquitin.
Figures
Similar articles
-
Allosteric regulation of deubiquitylase activity through ubiquitination.Front Mol Biosci. 2015 Feb 5;2:2. doi: 10.3389/fmolb.2015.00002. eCollection 2015. Front Mol Biosci. 2015. PMID: 25988170 Free PMC article.
-
Understanding the role of the Josephin domain in the PolyUb binding and cleavage properties of ataxin-3.PLoS One. 2010 Aug 26;5(8):e12430. doi: 10.1371/journal.pone.0012430. PLoS One. 2010. PMID: 20865150 Free PMC article.
-
Structural insights into the activity and regulation of human Josephin-2.J Struct Biol X. 2019 Aug 21;3:100011. doi: 10.1016/j.yjsbx.2019.100011. eCollection 2019 Jul-Sep. J Struct Biol X. 2019. PMID: 32647816 Free PMC article.
-
Regulation of proteolysis by human deubiquitinating enzymes.Biochim Biophys Acta. 2014 Jan;1843(1):114-28. doi: 10.1016/j.bbamcr.2013.06.027. Epub 2013 Jul 9. Biochim Biophys Acta. 2014. PMID: 23845989 Free PMC article. Review.
-
Chemical approaches for the preparation of ubiquitinated proteins via natural linkages.J Pept Sci. 2022 Mar;28(3):e3367. doi: 10.1002/psc.3367. Epub 2021 Sep 12. J Pept Sci. 2022. PMID: 34514672 Review.
Cited by
-
Chemical Protein Ubiquitylation with Preservation of the Native Cysteine Residues.Chembiochem. 2016 Jun 2;17(11):995-8. doi: 10.1002/cbic.201600042. Epub 2016 Apr 26. Chembiochem. 2016. PMID: 27113245 Free PMC article.
-
Lysine 117 on ataxin-3 modulates toxicity in Drosophila models of Spinocerebellar Ataxia Type 3.J Neurol Sci. 2023 Nov 15;454:120828. doi: 10.1016/j.jns.2023.120828. Epub 2023 Oct 5. J Neurol Sci. 2023. PMID: 37865002 Free PMC article.
-
The challenge of producing ubiquitinated proteins for structural studies.Cells. 2014 Jun 12;3(2):639-56. doi: 10.3390/cells3020639. Cells. 2014. PMID: 24926903 Free PMC article.
-
Allosteric regulation of deubiquitylase activity through ubiquitination.Front Mol Biosci. 2015 Feb 5;2:2. doi: 10.3389/fmolb.2015.00002. eCollection 2015. Front Mol Biosci. 2015. PMID: 25988170 Free PMC article.
-
Nonspecific yet decisive: Ubiquitination can affect the native-state dynamics of the modified protein.Protein Sci. 2015 Oct;24(10):1580-92. doi: 10.1002/pro.2688. Epub 2015 Jun 9. Protein Sci. 2015. PMID: 25970168 Free PMC article.
References
-
- Goldknopf I.L., Taylor C.W., Baum R.M., Yeoman L.C., Olson M.O., Prestayko A.W., Busch H. Isolation and characterization of protein A24, a “histone-like” non-histone chromosomal protein. J. Biol. Chem. 1975;250:7182–7187. - PubMed
-
- Sadowski M., Suryadinata R., Tan A.R., Roesley S.N., Sarcevic B. Protein monoubiquitination and polyubiquitination generate structural diversity to control distinct biological processes. IUBMB Life. 2012;64:136–142. - PubMed
-
- Komander D., Rape M. The ubiquitin code. Annu. Rev. Biochem. 2012;81:203–229. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
