

Intracellular glutathione pools are heterogeneously concentrated
- PMID: 24251119
- PMCID: PMC3830055
- DOI: 10.1016/j.redox.2013.10.005
Intracellular glutathione pools are heterogeneously concentrated
Abstract
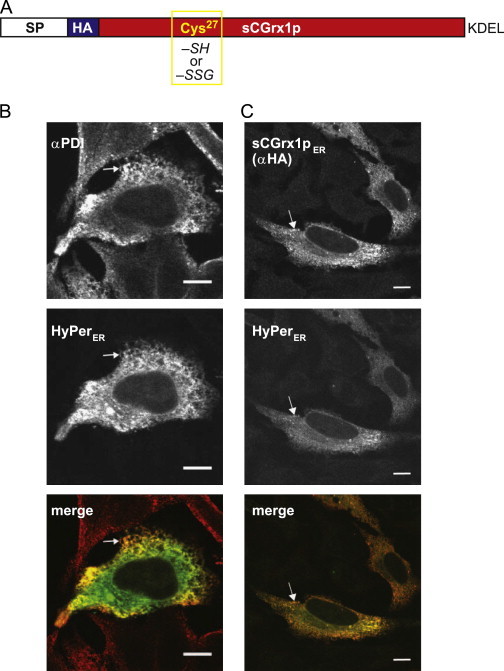
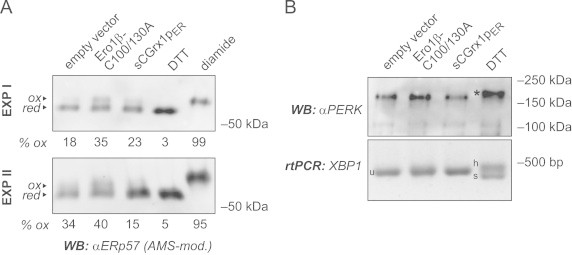
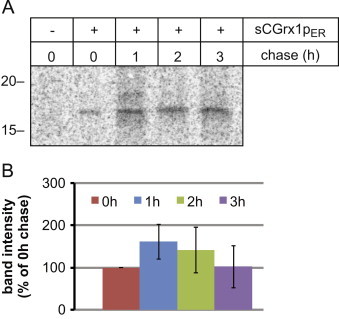
Glutathione is present in millimolar concentrations in the cell, but its relative distribution among cellular compartments remains elusive. We have chosen the endoplasmic reticulum (ER) as an example organelle to study compartment-specific glutathione levels. Using a glutaredoxin sensor (sCGrx1pER), which rapidly and specifically equilibrates with the reduced glutathione (GSH)-glutathione disulfide (GSSG) redox couple with known equilibrium constant, we showed that the [GSH]:[GSSG] ratio in the ER of intact HeLa cells is less than 7:1. Taking into consideration the previously determined value for [GSH](2):[GSSG] in the ER of 83 mM, this translates into a total glutathione concentration in the ER ([GStot]=[GSH]+2[GSSG]) of greater than 15 mM. Since the integrated, intracellular [GStot] was measured as ~7 mM, we conclude the existence of a [GStot] gradient across the ER membrane. A possible homeostatic mechanism by which cytosol-derived glutathione is trapped in the ER is discussed. We propose a high [GStot] as a distinguishing feature of the ER environment compared to the extracellular space.
Keywords: DTT, Dithiothreitol; EGSH, Half cell reduction potential of glutathione; ER, Endoplasmic reticulum; Endoplasmic reticulum; GSH, Reduced glutathione; GSSG, Glutathione disulfide; Glutaredoxin; Glutathione; NEM, N-ethylmaleimide; OxD, Percentage of oxidation; PDI, Protein disulfide isomerase; PERK, Double stranded RNA-activated protein kinase (PKR)-like ER kinase; RGS, [GSH]:[GSSG]; Redox Homeostasis; Redox compartmentalization; Redox, Reduction–oxidation; Reduction potential; TMM(PEG)12, Maleimide-activated polyethylene glycol; UPR, Unfolded protein response; XBP1, X-box binding protein 1; [GStot], Total glutathione concentration; sCGrx1p, C30S mutant of yeast glutaredoxin 1.
Figures


Similar articles
-
Oxidative protein folding and unfolded protein response elicit differing redox regulation in endoplasmic reticulum and cytosol of yeast.Free Radic Biol Med. 2012 May 1;52(9):2000-12. doi: 10.1016/j.freeradbiomed.2012.02.048. Epub 2012 Mar 8. Free Radic Biol Med. 2012. PMID: 22406321
-
Endoplasmic reticulum: reduced and oxidized glutathione revisited.J Cell Sci. 2013 Apr 1;126(Pt 7):1604-17. doi: 10.1242/jcs.117218. Epub 2013 Feb 19. J Cell Sci. 2013. PMID: 23424194
-
Assessment of endoplasmic reticulum glutathione redox status is confounded by extensive ex vivo oxidation.Antioxid Redox Signal. 2008 May;10(5):963-72. doi: 10.1089/ars.2007.1869. Antioxid Redox Signal. 2008. PMID: 18205546 Free PMC article.
-
Molecular Mechanisms of Glutaredoxin Enzymes: Versatile Hubs for Thiol-Disulfide Exchange between Protein Thiols and Glutathione.J Mol Biol. 2019 Jan 18;431(2):158-177. doi: 10.1016/j.jmb.2018.12.006. Epub 2018 Dec 12. J Mol Biol. 2019. PMID: 30552876 Review.
-
The glutathione system and the related thiol network in Caenorhabditis elegans.Redox Biol. 2019 Jun;24:101171. doi: 10.1016/j.redox.2019.101171. Epub 2019 Mar 16. Redox Biol. 2019. PMID: 30901603 Free PMC article. Review.
Cited by
-
S-glutathionylation, friend or foe in cardiovascular health and disease.Redox Biol. 2020 Oct;37:101693. doi: 10.1016/j.redox.2020.101693. Epub 2020 Aug 22. Redox Biol. 2020. PMID: 32912836 Free PMC article. Review.
-
Effect of Low Concentration of Nitroxides on SH-SY5Y Cells Transfected with the Tau Protein.Int J Mol Sci. 2023 Nov 23;24(23):16675. doi: 10.3390/ijms242316675. Int J Mol Sci. 2023. PMID: 38069000 Free PMC article.
-
S-Glutathionylation-Controlled Apoptosis of Lung Epithelial Cells; Potential Implications for Lung Fibrosis.Antioxidants (Basel). 2022 Sep 10;11(9):1789. doi: 10.3390/antiox11091789. Antioxidants (Basel). 2022. PMID: 36139863 Free PMC article. Review.
-
Modeling Reactivity to Biological Macromolecules with a Deep Multitask Network.ACS Cent Sci. 2016 Aug 24;2(8):529-37. doi: 10.1021/acscentsci.6b00162. Epub 2016 Jul 29. ACS Cent Sci. 2016. PMID: 27610414 Free PMC article.
-
Preserving Single Cells in Space and Time for Analytical Assays.Trends Analyt Chem. 2020 Jan;122:115723. doi: 10.1016/j.trac.2019.115723. Epub 2019 Nov 7. Trends Analyt Chem. 2020. PMID: 32153309 Free PMC article.
References
-
- Deponte M. Glutathione catalysis and the reaction mechanisms of glutathione-dependent enzymes. Biochim. Biophys. Acta. 2013;1830:3217–3266. - PubMed
-
- Bachhawat A.K., Thakur A., Kaur J., Zulkifli M. Glutathione transporters. Biochim. Biophys. Acta. 2013;1830:3154–3164. - PubMed
-
- Appenzeller-Herzog C. Glutathione- and non-glutathione-based oxidant control in the endoplasmic reticulum. J. Cell Sci. 2011;124:847–855. - PubMed
-
- Ramming T., Appenzeller-Herzog C. The physiological functions of mammalian endoplasmic oxidoreductin 1: on disulfides and more. Antioxid. Redox Signal. 2012;16:1109–1118. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
