

A quantitative model of amphetamine action on the 5-HT transporter
- PMID: 24251585
- PMCID: PMC3925039
- DOI: 10.1111/bph.12520
A quantitative model of amphetamine action on the 5-HT transporter
Abstract
Background and purpose: Amphetamines bind to the plasmalemmal transporters for the monoamines dopamine (DAT), noradrenaline (NET) and 5-HT (SERT); influx of amphetamine leads to efflux of substrates. Various models have been proposed to account for this amphetamine-induced reverse transport in mechanistic terms. A most notable example is the molecular stent hypothesis, which posits a special amphetamine-induced conformation that is not likely in alternative access models of transport. The current study was designed to evaluate the explanatory power of these models and the molecular stent hypothesis.
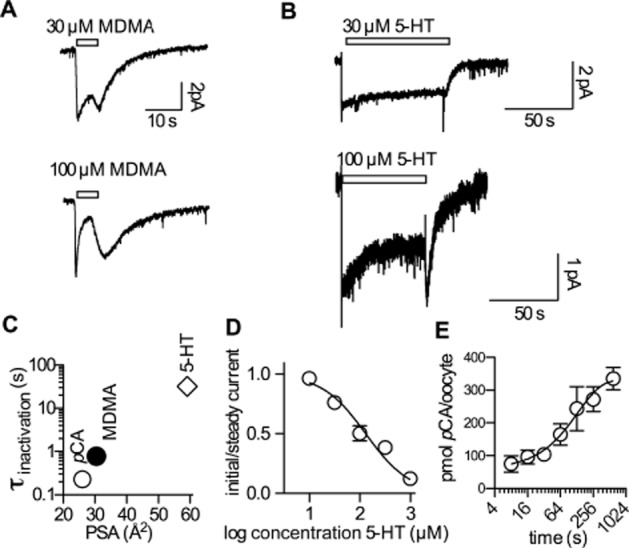
Experimental approach: Xenopus laevis oocytes and HEK293 cells expressing human (h) SERT were voltage-clamped and exposed to 5-HT, p-chloroamphetamine (pCA) or methylenedioxyamphetamine (MDMA).
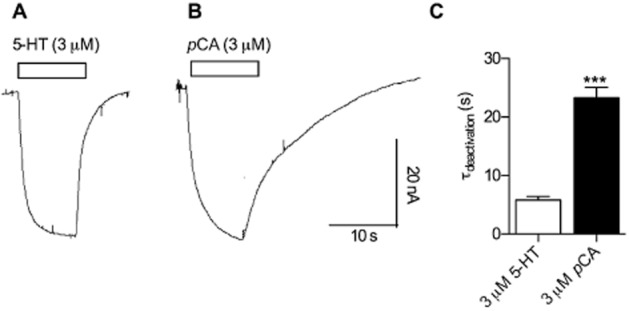
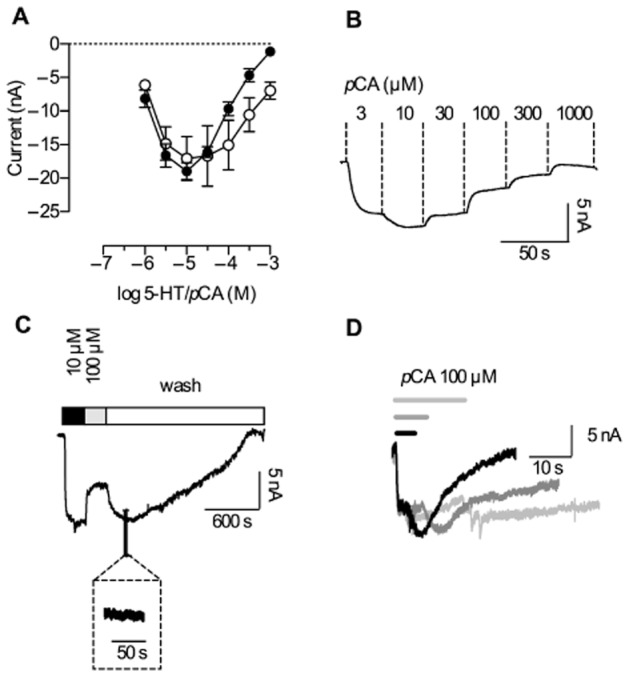
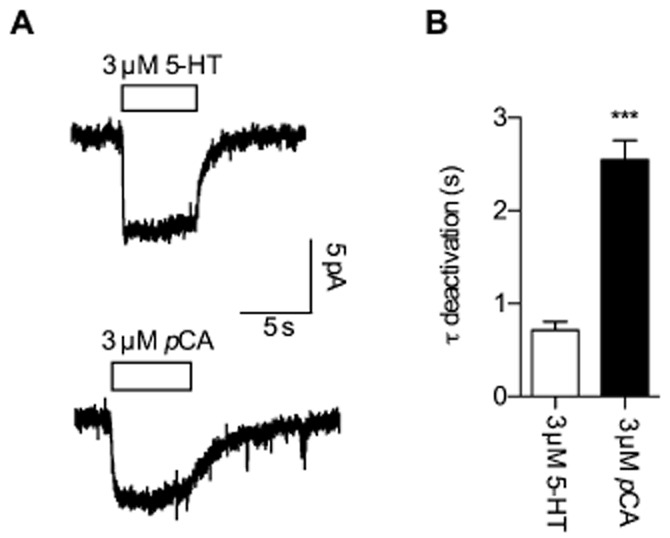
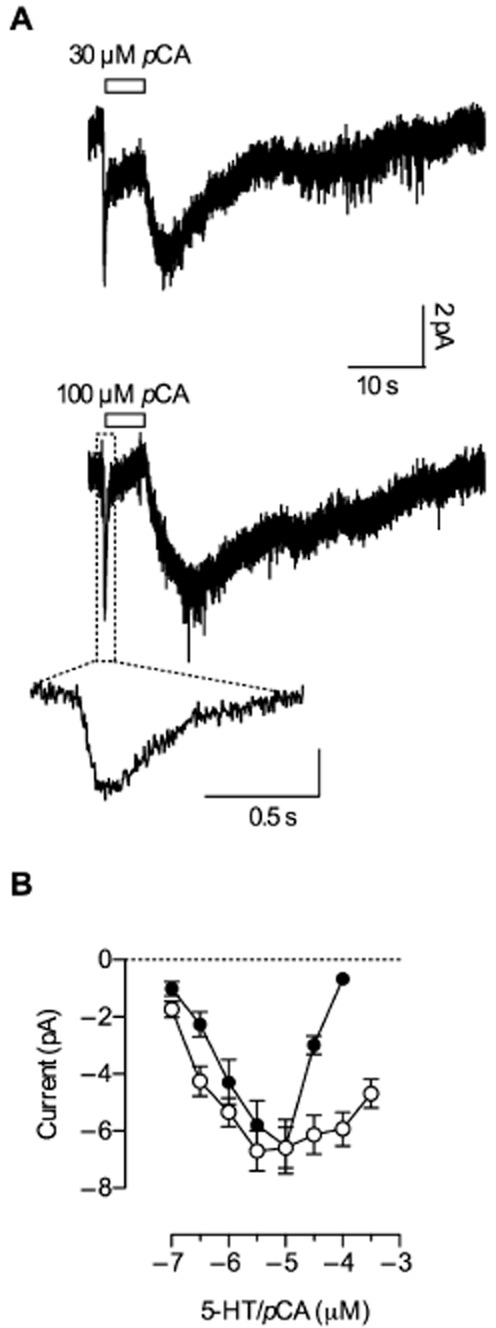
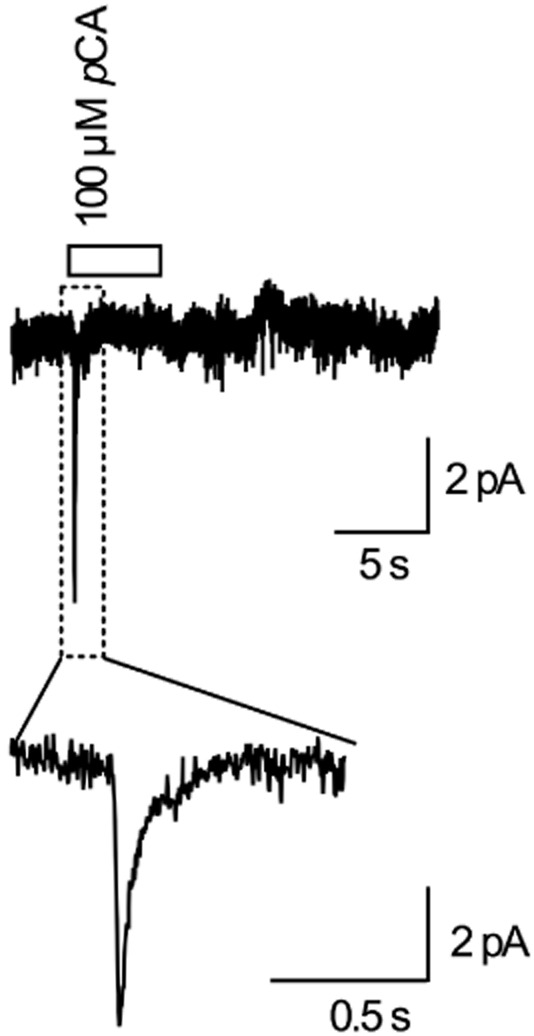
Key results: In contrast to the currents induced by 5-HT, pCA-triggered currents through SERT decayed slowly in Xenopus laevis oocytes once the agonist was removed (consistent with the molecular stent hypothesis). However, when SERT was expressed in HEK293 cells, currents induced by 3 or 100 μM pCA decayed 10 or 100 times faster, respectively, after pCA removal.
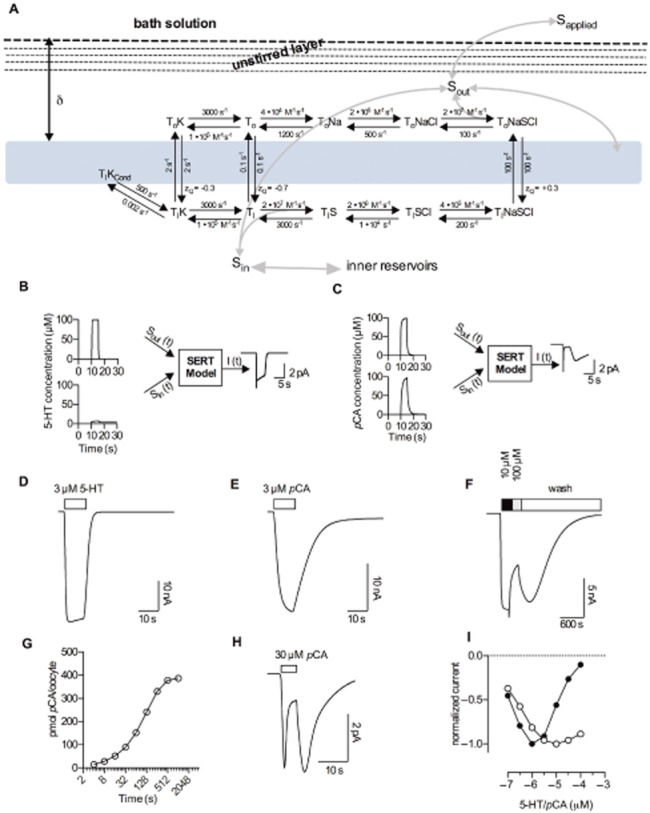
Conclusions and implications: This discrepancy in decay rates is inconsistent with the molecular stent hypothesis. In contrast, a multistate version of the alternative access model accounts for all the observations and reproduces the kinetic parameters extracted from the electrophysiological recordings. A crucial feature that explains the action of amphetamines is their lipophilic nature, which allows for rapid diffusion through the membrane.
Keywords: SERT; amphetamine; currents; diffusion.
© 2013 The Authors. British Journal of Pharmacology published by John Wiley &. Sons Ltd on behalf of The British Pharmacological Society.
Figures
Comment in
-
The conservative view: is it necessary to implant a stent into the dopamine transporter?Br J Pharmacol. 2015 Oct;172(19):4775-8. doi: 10.1111/bph.12766. Br J Pharmacol. 2015. PMID: 24824446 Free PMC article.
-
Comments on 'A quantitative model of amphetamine action on the serotonin transporter', by Sandtner et al., Br J Pharmacol 171: 1007-1018.Br J Pharmacol. 2015 Oct;172(19):4772-4. doi: 10.1111/bph.12767. Br J Pharmacol. 2015. PMID: 24824549 Free PMC article.
References
-
- Baldwin RL. The new view of hydrophobic free energy. FEBS Lett. 2013;587:1062–1066. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
