

Stabilization of the dimeric birch pollen allergen Bet v 1 impacts its immunological properties
- PMID: 24253036
- PMCID: PMC3879576
- DOI: 10.1074/jbc.M113.518795
Stabilization of the dimeric birch pollen allergen Bet v 1 impacts its immunological properties
Abstract
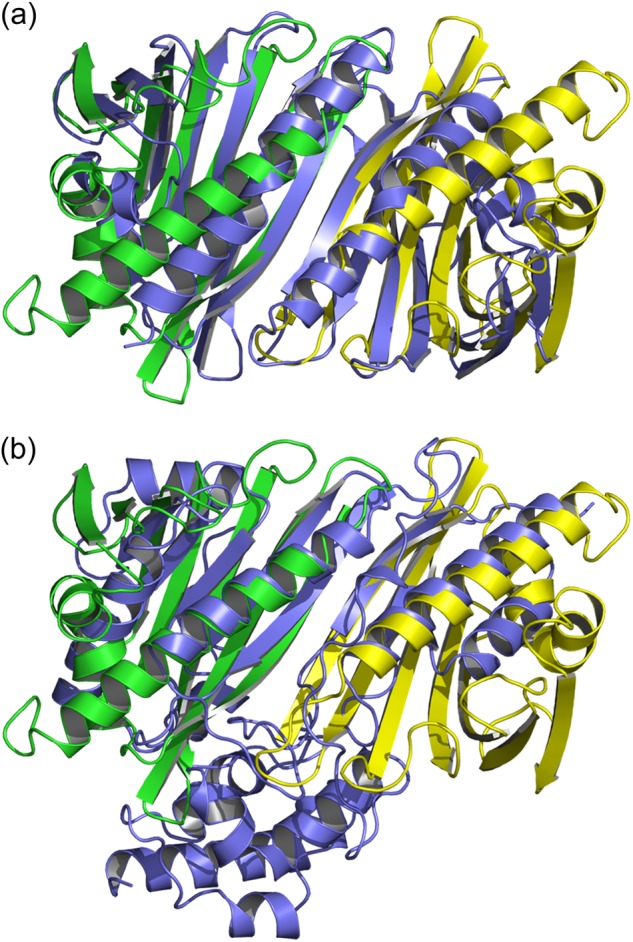
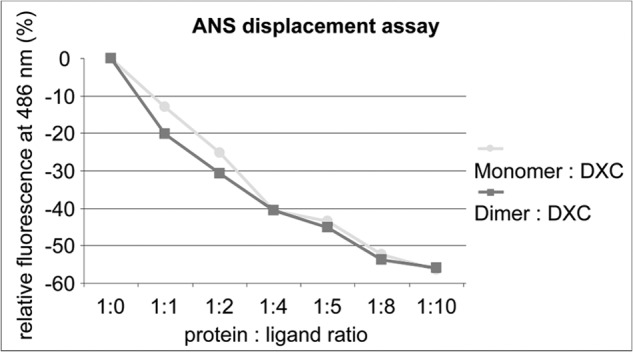
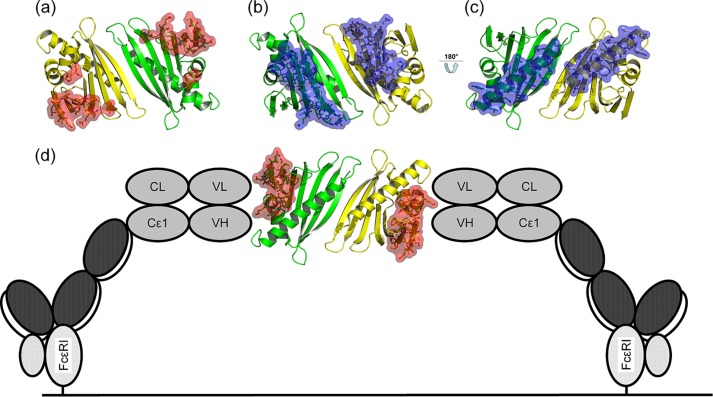
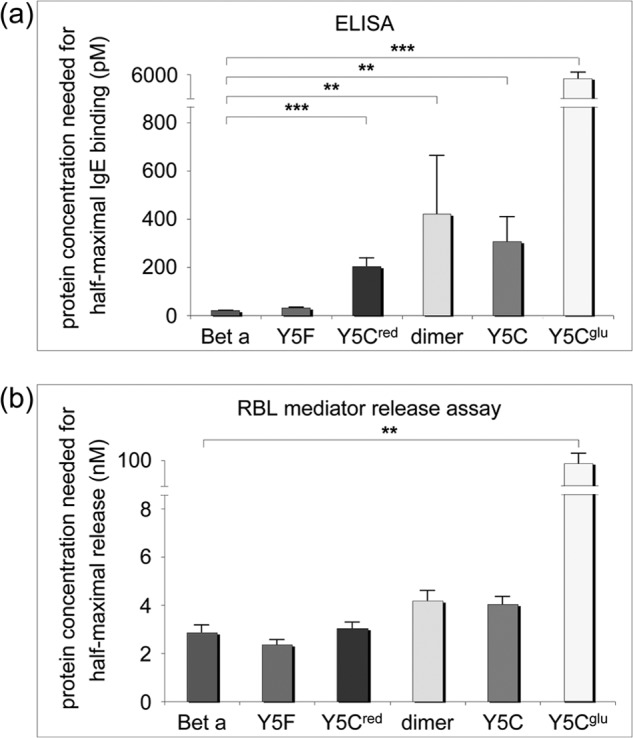
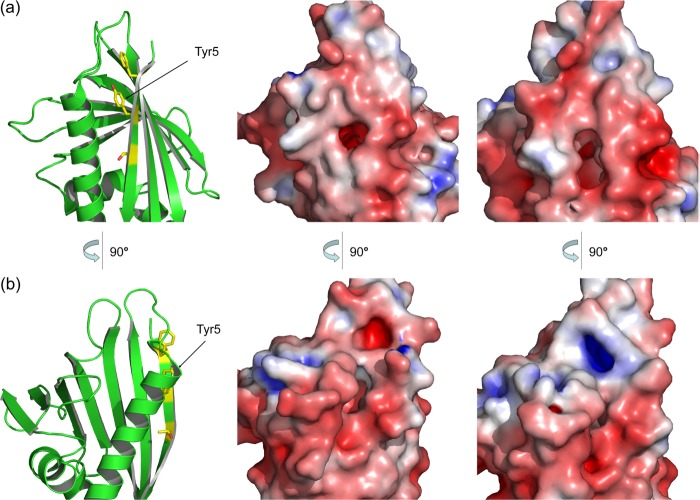
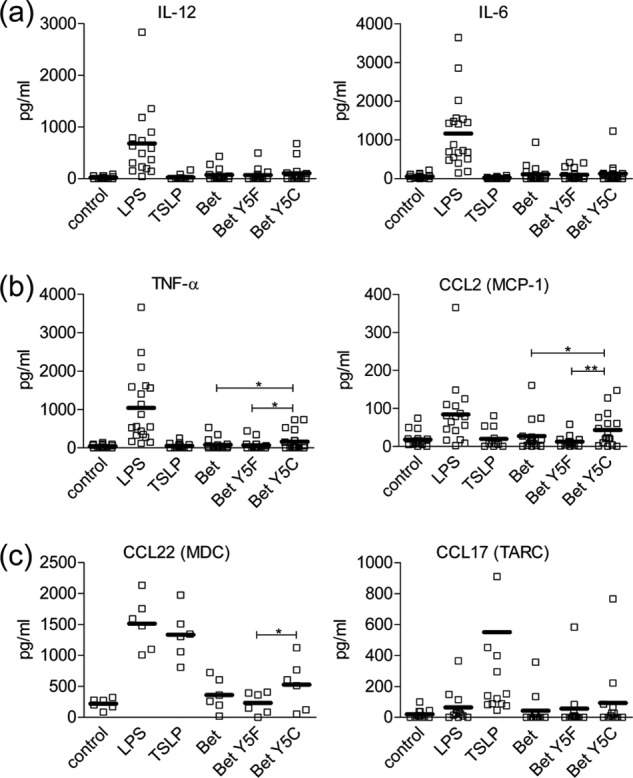
Many allergens share several biophysical characteristics, including the capability to undergo oligomerization. The dimerization mechanism in Bet v 1 and its allergenic properties are so far poorly understood. Here, we report crystal structures of dimeric Bet v 1, revealing a noncanonical incorporation of cysteine at position 5 instead of genetically encoded tyrosine. Cysteine polysulfide bridging stabilized different dimeric assemblies, depending on the polysulfide linker length. These dimers represent quaternary arrangements that are frequently observed in related proteins, reflecting their prevalence in unmodified Bet v 1. These conclusions were corroborated by characteristic immunologic properties of monomeric and dimeric allergen variants. Hereby, residue 5 could be identified as an allergenic hot spot in Bet v 1. The presented results refine fundamental principles in protein chemistry and emphasize the importance of protein modifications in understanding the molecular basis of allergenicity.
Keywords: Allergen; Crystal Structure; Dimerization; Mass Spectrometry (MS); Noncanonical Amino Acid Incorporation; Polysulfide Linking; Position-specific Alteration of Genetic Code; Post-translational Modification; Protein Assembly.
Figures
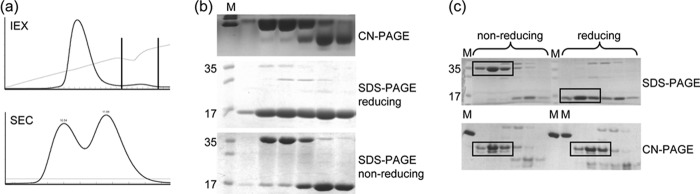
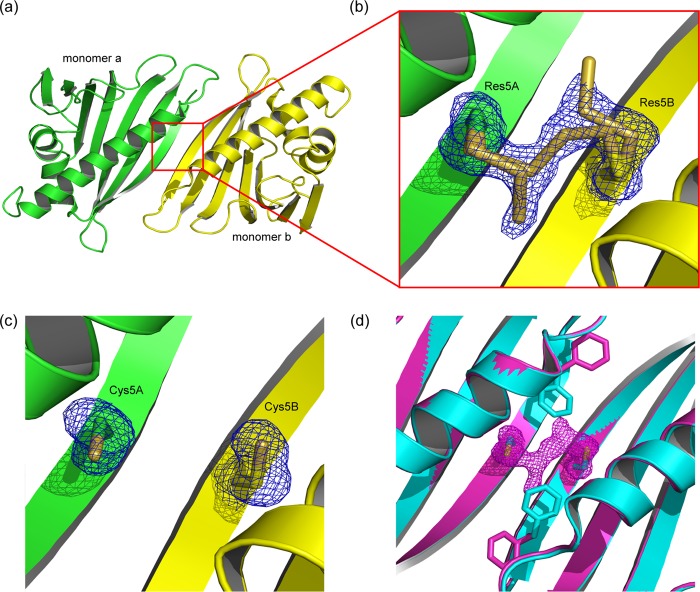
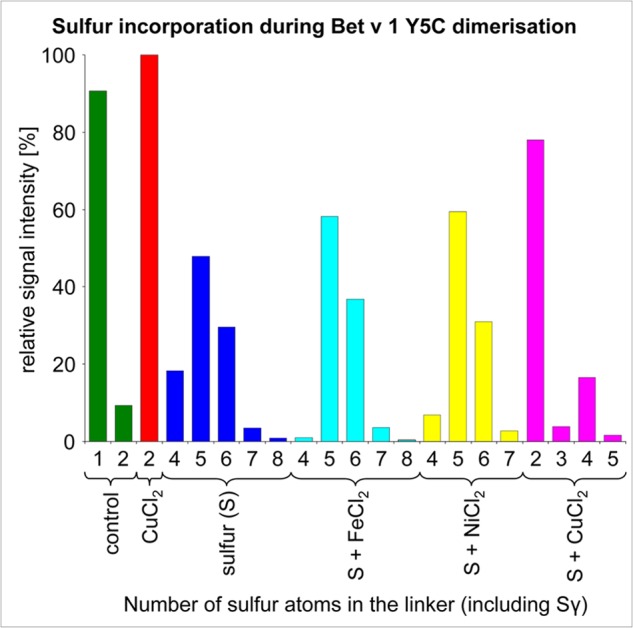
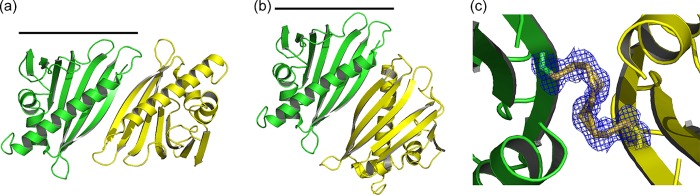
References
-
- Shakib F., Ghaemmaghami A. M., Sewell H. F. (2008) The molecular basis of allergenicity. Trends Immunol. 29, 633–642 - PubMed
-
- Lascombe M. B., Grégoire C., Poncet P., Tavares G. A., Rosinski-Chupin I., Rabillon J., Goubran-Botros H., Mazié J. C., David B., Alzari P. M. (2000) Crystal structure of the allergen Equ c 1. A dimeric lipocalin with restricted IgE-reactive epitopes. J. Biol. Chem. 275, 21572–21577 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
