

Platelet-targeted gene therapy with human factor VIII establishes haemostasis in dogs with haemophilia A
- PMID: 24253479
- PMCID: PMC3868233
- DOI: 10.1038/ncomms3773
Platelet-targeted gene therapy with human factor VIII establishes haemostasis in dogs with haemophilia A
Abstract
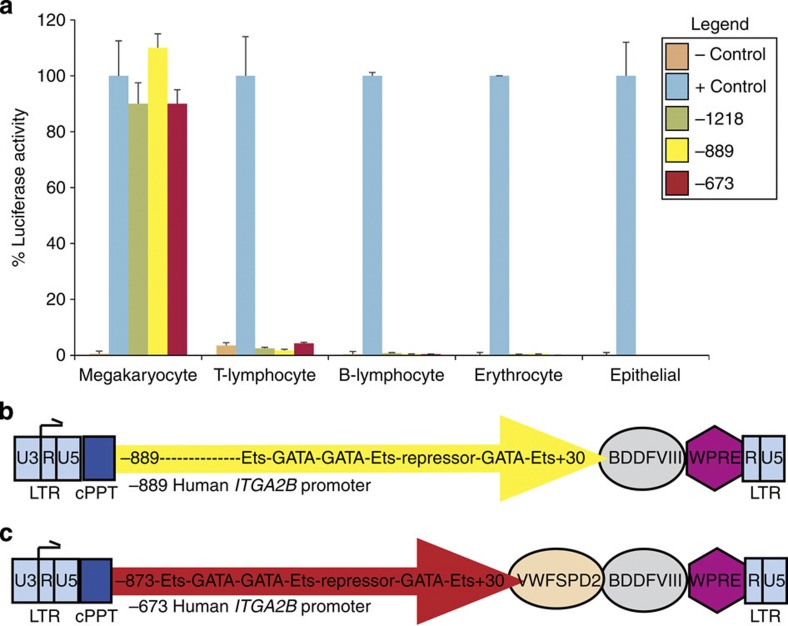
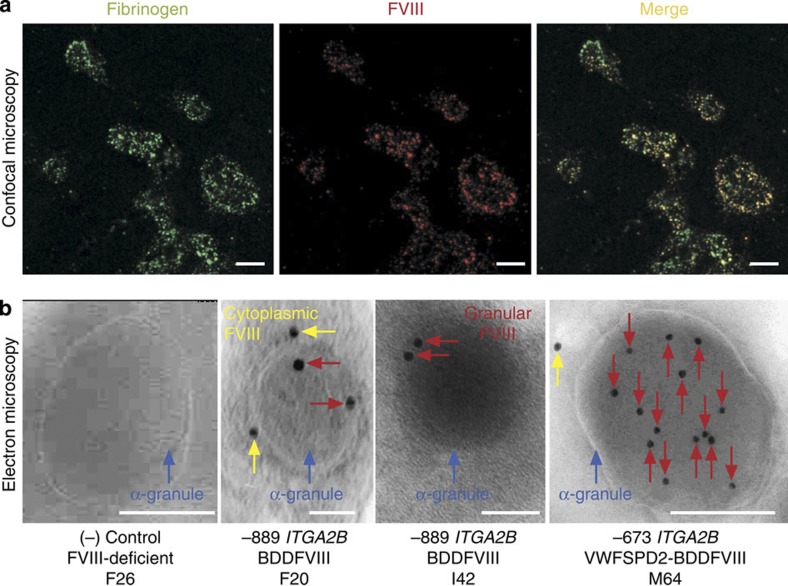
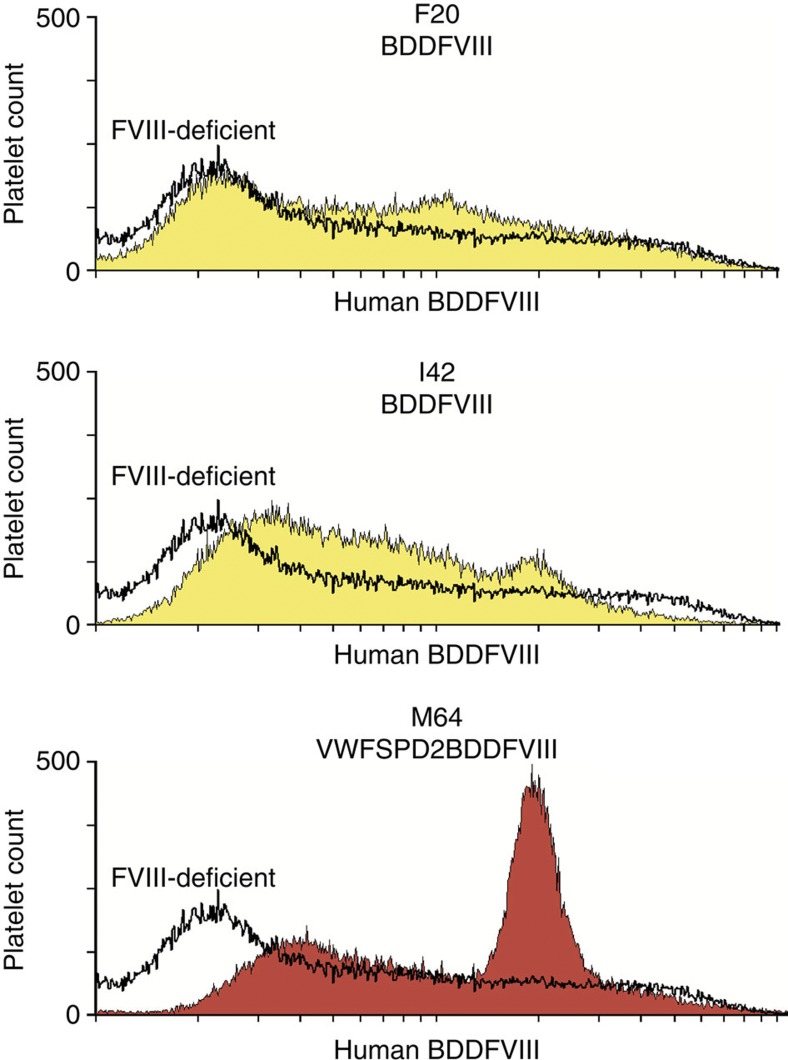
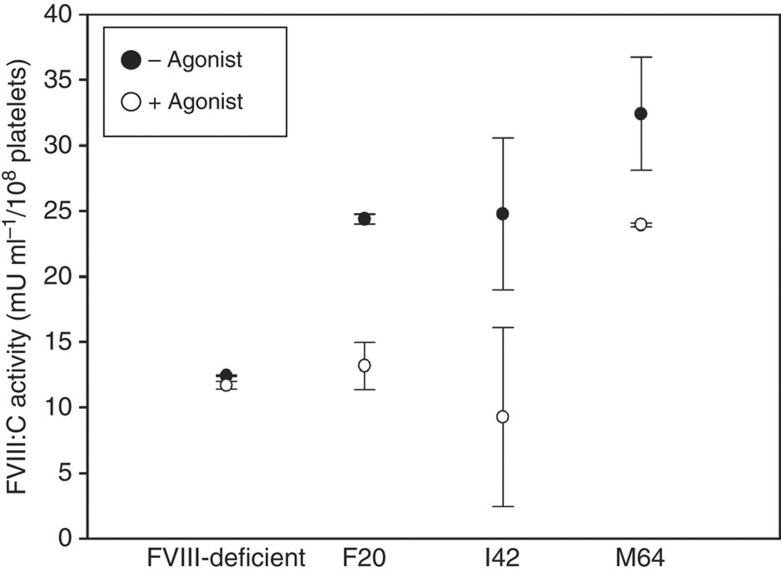
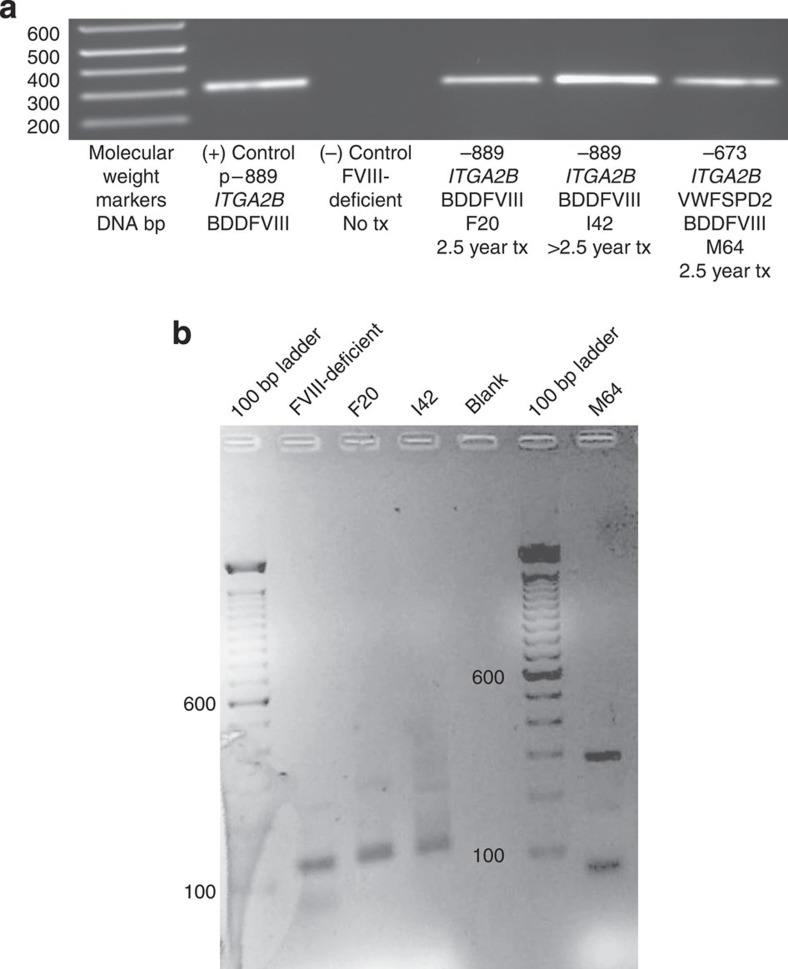
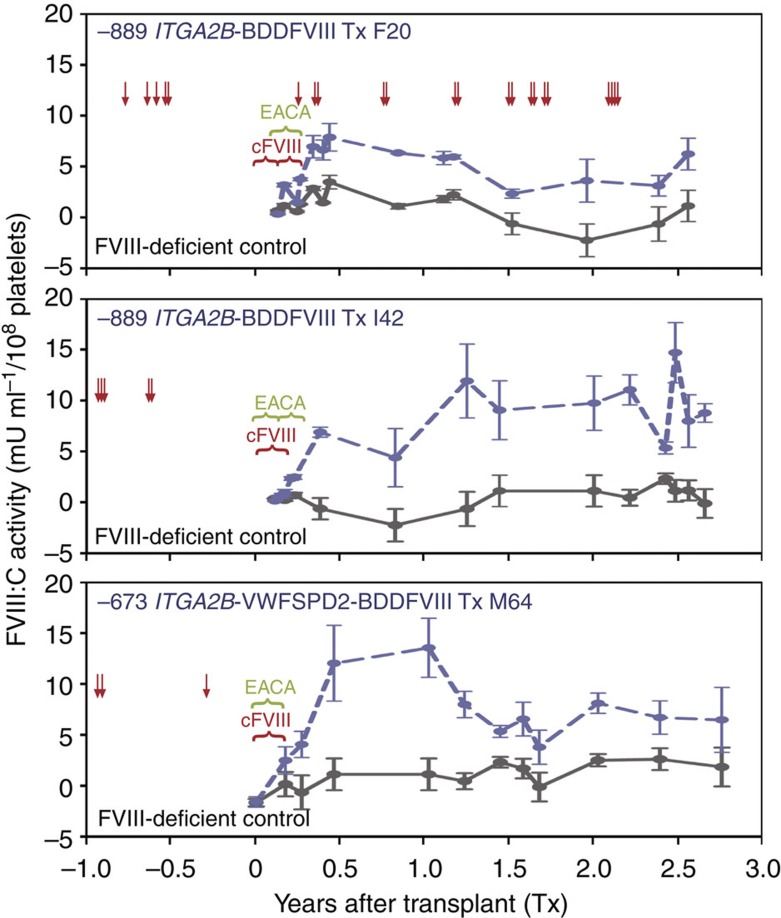
It is essential to improve therapies for controlling excessive bleeding in patients with haemorrhagic disorders. As activated blood platelets mediate the primary response to vascular injury, we hypothesize that storage of coagulation Factor VIII within platelets may provide a locally inducible treatment to maintain haemostasis for haemophilia A. Here we show that haematopoietic stem cell gene therapy can prevent the occurrence of severe bleeding episodes in dogs with haemophilia A for at least 2.5 years after transplantation. We employ a clinically relevant strategy based on a lentiviral vector encoding the ITGA2B gene promoter, which drives platelet-specific expression of human FVIII permitting storage and release of FVIII from activated platelets. One animal receives a hybrid molecule of FVIII fused to the von Willebrand Factor propeptide-D2 domain that traffics FVIII more effectively into α-granules. The absence of inhibitory antibodies to platelet-derived FVIII indicates that this approach may have benefit in patients who reject FVIII replacement therapies. Thus, platelet FVIII may provide effective long-term control of bleeding in patients with haemophilia A.
Conflict of interest statement
David A. Wilcox and Sandra L. Haberichter have applied for a US Provisional Patent Application (serial no. 61/717,951) entitled “Platelet Targeted Treatment” for the therapy described within this manuscript. All other authors declare no competing interests.
Figures
References
-
- Leslie M. Cell biology. Beyond clotting: the powers of platelets. Science 328, 562–564 (2010). - PubMed
-
- Stonebraker J. S., Bolton-Maggs P. H., Soucie J. M., Walker I. & Brooker M. A study of variations in the reported haemophilia A prevalence around the world. Haemophilia 16, 20–32 (2010). - PubMed
-
- Berntorp E. & Shapiro A. D. Modern haemophilia care. Lancet 379, 1447–1456 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
