

Identification of RNase L-dependent, 3'-end-modified, viral small RNAs in Sindbis virus-infected mammalian cells
- PMID: 24255120
- PMCID: PMC3870239
- DOI: 10.1128/mBio.00698-13
Identification of RNase L-dependent, 3'-end-modified, viral small RNAs in Sindbis virus-infected mammalian cells
Abstract
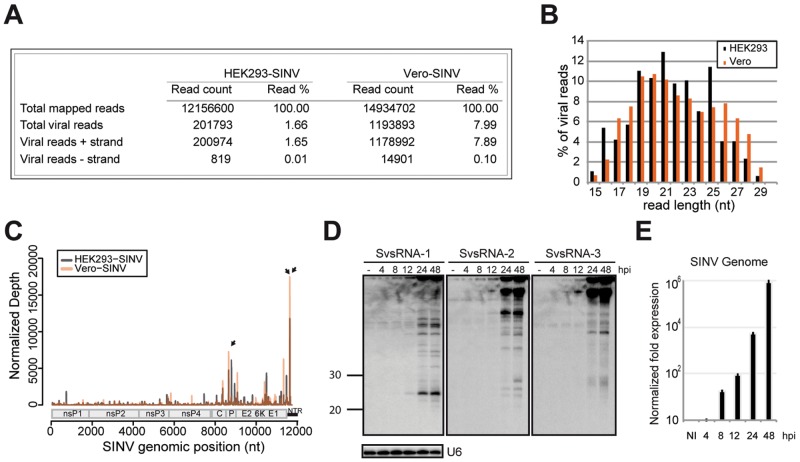
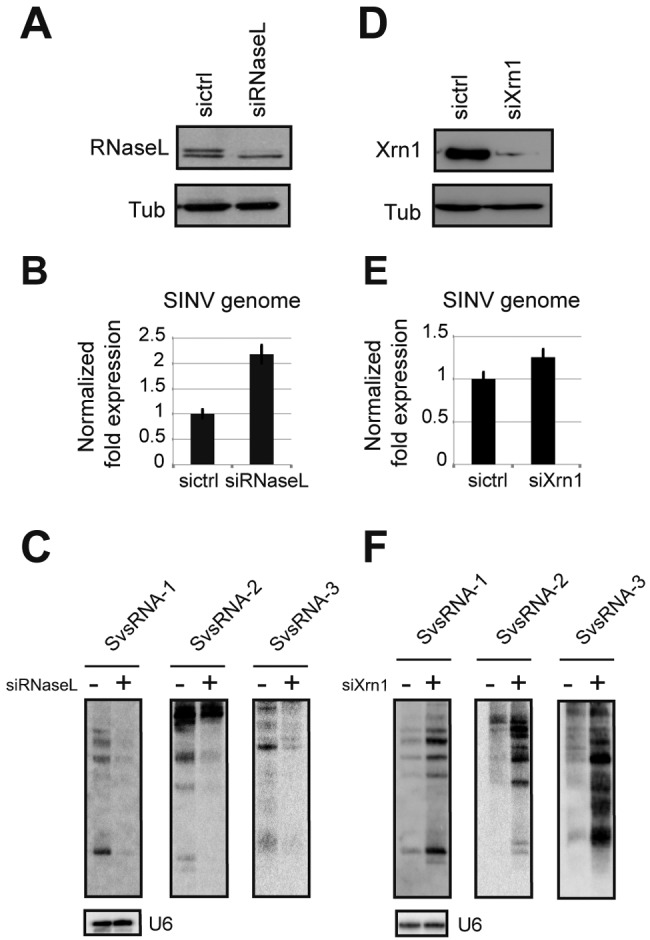
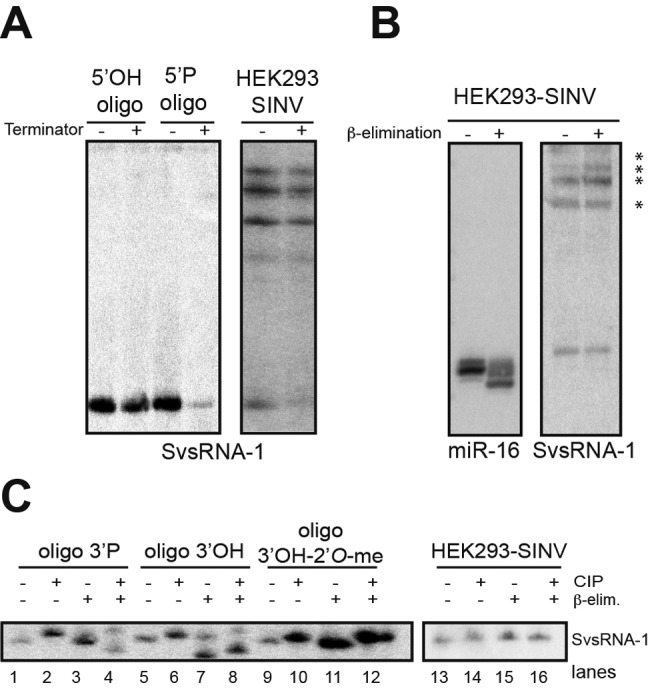
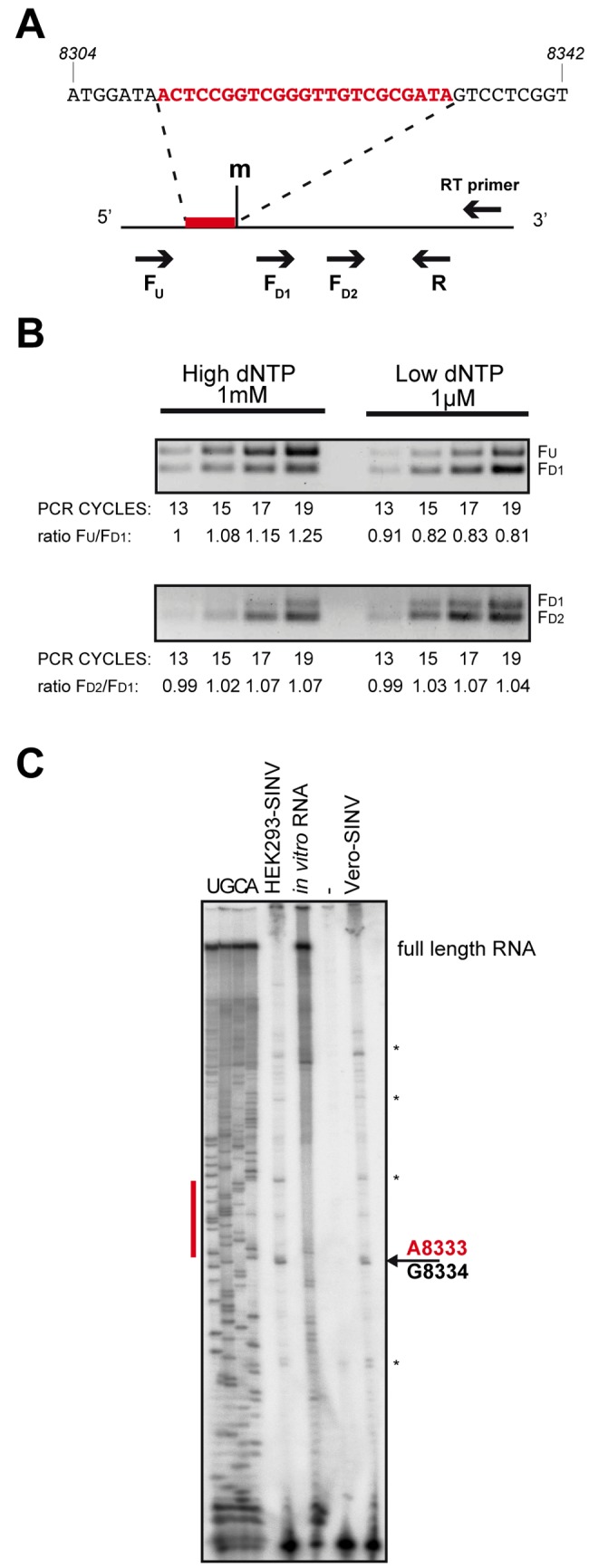
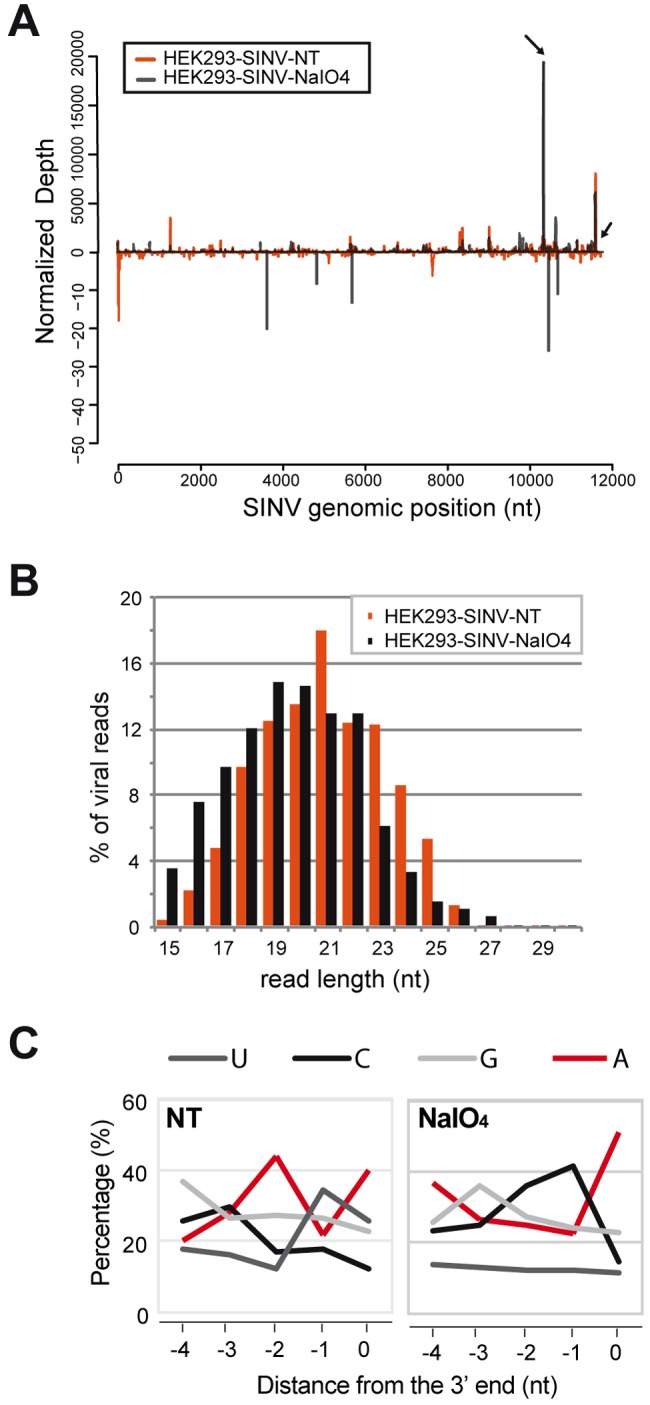
Small RNAs play a critical role in host-pathogen interaction. Indeed, small RNA-mediated silencing or RNA interference (RNAi) is one of the earliest forms of antiviral immunity. Although it represents the main defense system against viruses in many organisms, the antiviral role of RNAi has not been clearly proven in higher vertebrates. However, it is well established that their response to viral infection relies on the recognition of viral RNAs by host pattern recognition receptors (PRRs) to trigger activation of the interferon pathway. In the present work, we report the existence of a novel small noncoding RNA population produced in mammalian cells upon RNA virus infection. Using Sindbis virus (SINV) as a prototypic arbovirus model, we profiled the small RNA population of infected cells in both human and African green monkey cell lines. Here, we provide evidence for the presence of discrete small RNAs of viral origin that are not associated with the RNA-induced silencing complex (RISC), that are highly expressed and detected by Northern blot analysis, and that accumulate as 21- to 28-nucleotide (nt) species during infection. We report that the cellular antiviral endoribonuclease RNase L cleaves the viral genome, producing in turn the small RNAs. Surprisingly, we uncovered the presence of a modification on the 3'-end nucleotide of SINV-derived viral small RNAs (SvsRNAs) that might be at the origin of their stability. Altogether, our findings show that stable modified small viral RNAs could represent a novel way to modulate host-virus interaction upon SINV infection.
Importance: In a continuous arms race, viruses have to deal with host antiviral responses in order to successfully establish an infection. In mammalian cells, the host defense mechanism relies on the recognition of viral RNAs, resulting in the activation of type I interferons (IFNs). In turn, the expression of many interferon-stimulated genes (ISGs) is induced to inhibit viral replication. Here we report that the cytoplasmic, interferon-induced, cellular endoribonuclease RNase L is involved in the accumulation of a novel small RNA population of viral origin. These small RNAs are produced upon SINV infection of mammalian cells and are stabilized by a 3'-end modification. Altogether, our findings indicate that in our system RNA silencing is not active against Sindbis virus (SINV) and might open the way to a better understanding of the antiviral response mediated by a novel class of small RNAs.
Figures
Similar articles
-
The evolving world of small RNAs from RNA viruses.Wiley Interdiscip Rev RNA. 2016 Sep;7(5):575-88. doi: 10.1002/wrna.1351. Epub 2016 Apr 5. Wiley Interdiscip Rev RNA. 2016. PMID: 27046163 Free PMC article. Review.
-
TMEΜ45B Interacts with Sindbis Virus Nsp1 and Nsp4 and Inhibits Viral Replication.J Virol. 2022 Sep 14;96(17):e0091922. doi: 10.1128/jvi.00919-22. Epub 2022 Aug 8. J Virol. 2022. PMID: 35938871 Free PMC article.
-
RNase L Amplifies Interferon Signaling by Inducing Protein Kinase R-Mediated Antiviral Stress Granules.J Virol. 2020 Jun 16;94(13):e00205-20. doi: 10.1128/JVI.00205-20. Print 2020 Jun 16. J Virol. 2020. PMID: 32295917 Free PMC article.
-
Activation of RNase L in Egyptian Rousette Bat-Derived RoNi/7 Cells Is Dependent Primarily on OAS3 and Independent of MAVS Signaling.mBio. 2019 Nov 12;10(6):e02414-19. doi: 10.1128/mBio.02414-19. mBio. 2019. PMID: 31719180 Free PMC article.
-
RNA-based viral immunity initiated by the Dicer family of host immune receptors.Immunol Rev. 2009 Jan;227(1):176-88. doi: 10.1111/j.1600-065X.2008.00722.x. Immunol Rev. 2009. PMID: 19120484 Free PMC article. Review.
Cited by
-
Production of functional small interfering RNAs by an amino-terminal deletion mutant of human Dicer.Proc Natl Acad Sci U S A. 2015 Dec 15;112(50):E6945-54. doi: 10.1073/pnas.1513421112. Epub 2015 Nov 30. Proc Natl Acad Sci U S A. 2015. PMID: 26621737 Free PMC article.
-
The RIG-I-like receptor LGP2 inhibits Dicer-dependent processing of long double-stranded RNA and blocks RNA interference in mammalian cells.EMBO J. 2018 Feb 15;37(4):e97479. doi: 10.15252/embj.201797479. Epub 2018 Jan 19. EMBO J. 2018. PMID: 29351913 Free PMC article.
-
Deletion of Cytoplasmic Double-Stranded RNA Sensors Does Not Uncover Viral Small Interfering RNA Production in Human Cells.mSphere. 2017 Aug 16;2(4):e00333-17. doi: 10.1128/mSphere.00333-17. eCollection 2017 Jul-Aug. mSphere. 2017. PMID: 28815217 Free PMC article.
-
Functional Mapping of AGO-Associated Zika Virus-Derived Small Interfering RNAs in Neural Stem Cells.Front Cell Infect Microbiol. 2021 Feb 25;11:628887. doi: 10.3389/fcimb.2021.628887. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 33718276 Free PMC article.
-
The evolving world of small RNAs from RNA viruses.Wiley Interdiscip Rev RNA. 2016 Sep;7(5):575-88. doi: 10.1002/wrna.1351. Epub 2016 Apr 5. Wiley Interdiscip Rev RNA. 2016. PMID: 27046163 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
