

β-lactoglobulin's conformational requirements for ligand binding at the calyx and the dimer interphase: a flexible docking study
- PMID: 24255705
- PMCID: PMC3821863
- DOI: 10.1371/journal.pone.0079530
β-lactoglobulin's conformational requirements for ligand binding at the calyx and the dimer interphase: a flexible docking study
Abstract
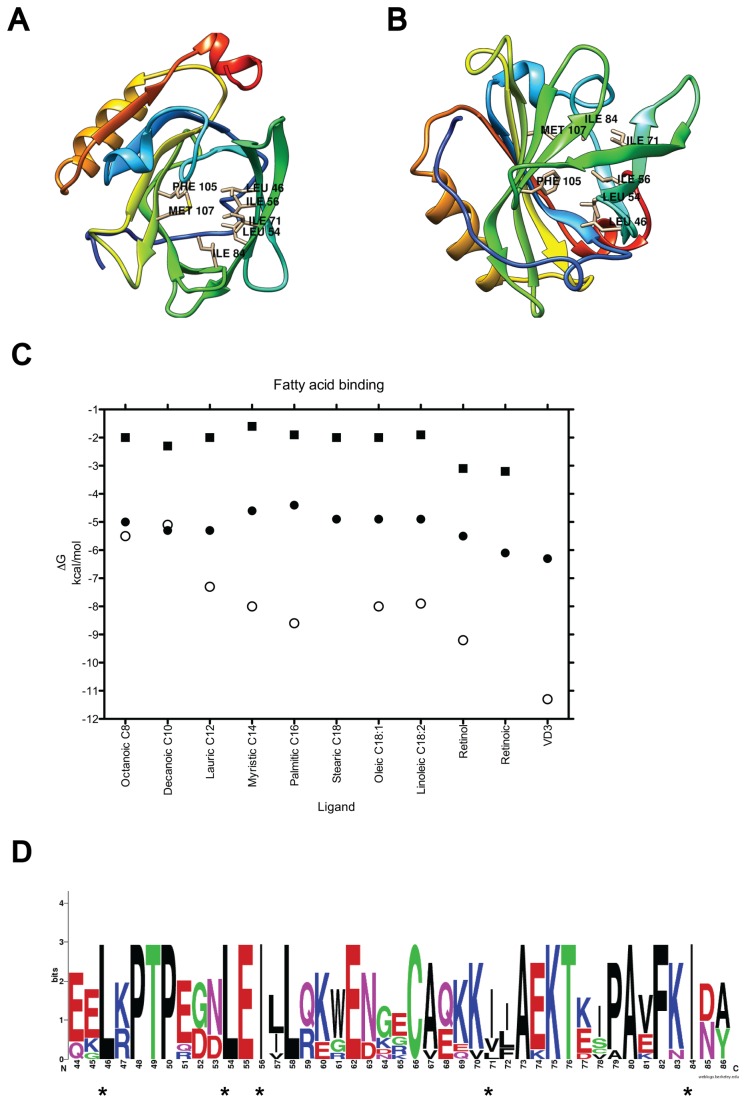
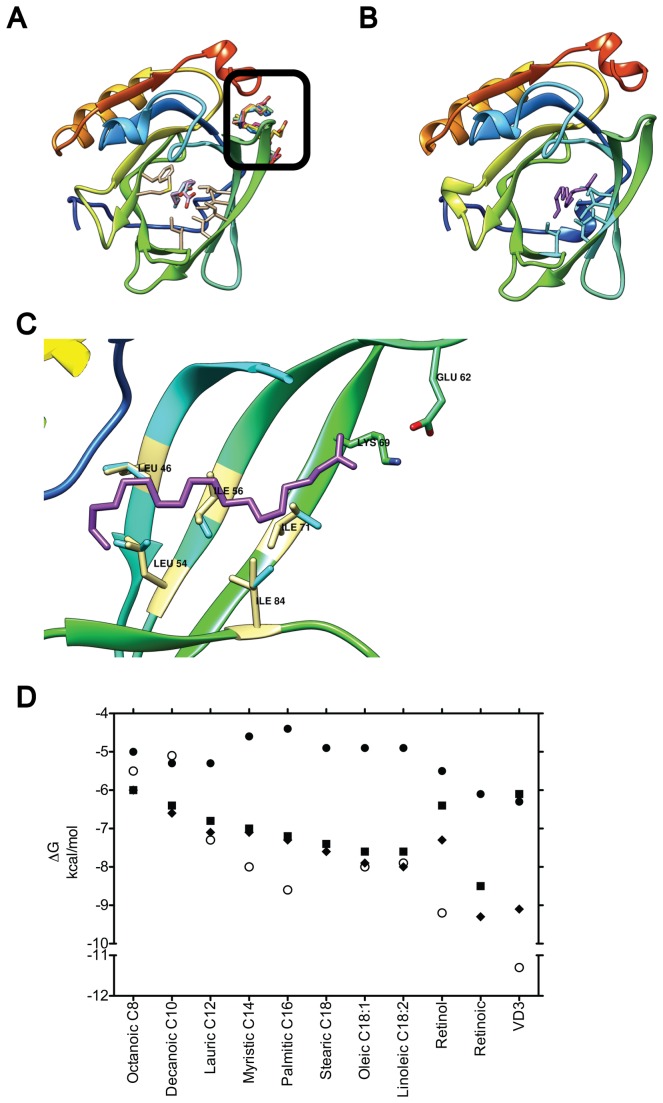
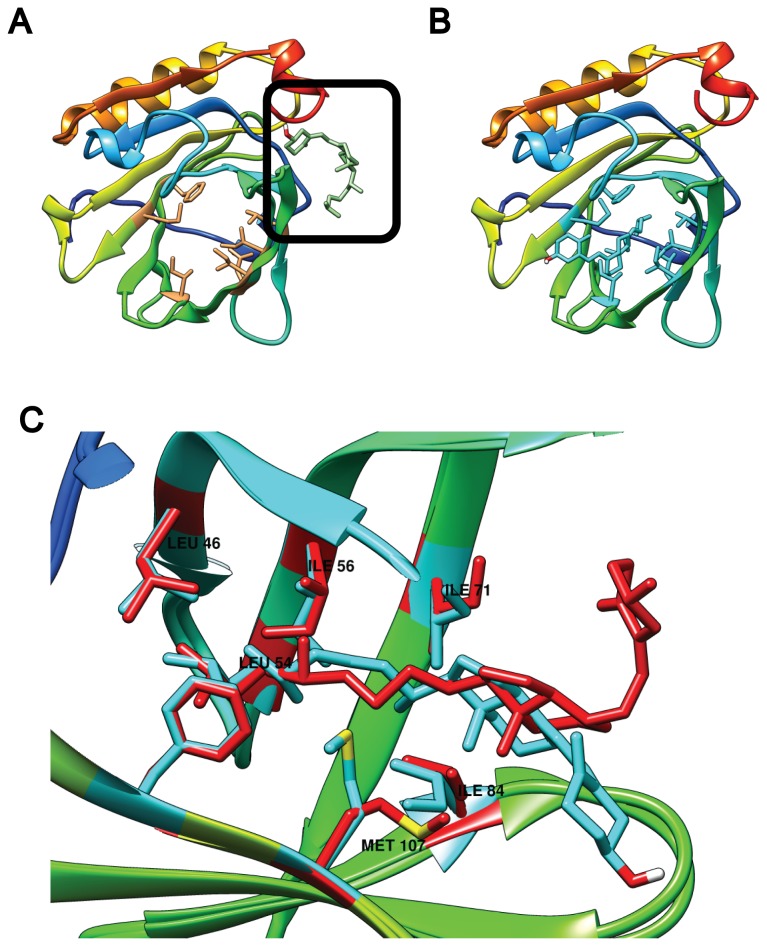
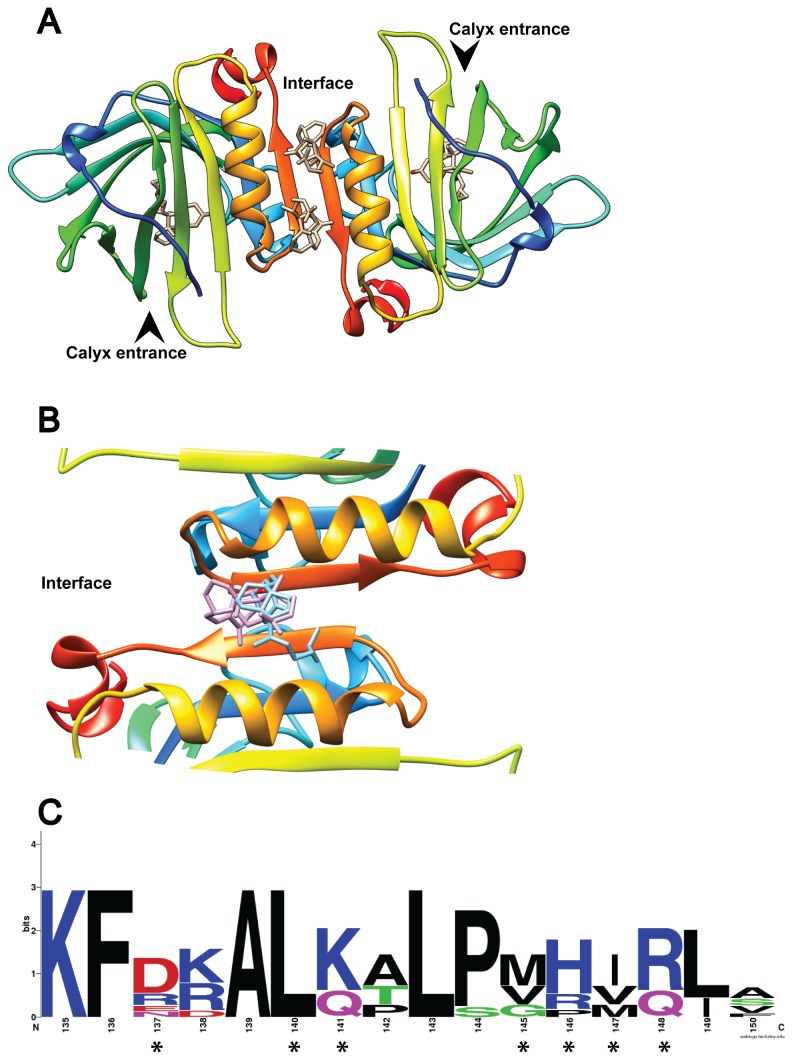
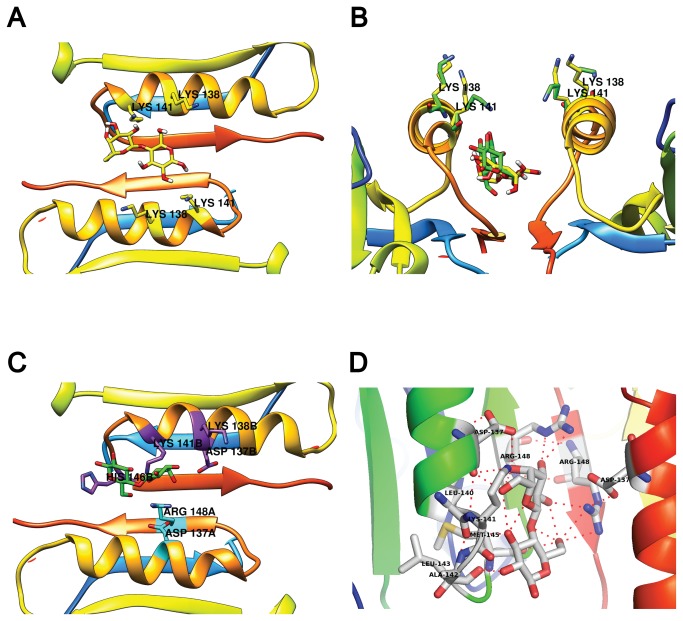
β-lactoglobulin (BLG) is an abundant milk protein relevant for industry and biotechnology, due significantly to its ability to bind a wide range of polar and apolar ligands. While hydrophobic ligand sites are known, sites for hydrophilic ligands such as the prevalent milk sugar, lactose, remain undetermined. Through the use of molecular docking we first, analyzed the known fatty acid binding sites in order to dissect their atomistic determinants and second, predicted the interaction sites for lactose with monomeric and dimeric BLG. We validated our approach against BLG structures co-crystallized with ligands and report a computational setup with a reduced number of flexible residues that is able to reproduce experimental results with high precision. Blind dockings with and without flexible side chains on BLG showed that: i) 13 experimentally-determined ligands fit the calyx requiring minimal movement of up to 7 residues out of the 23 that constitute this binding site. ii) Lactose does not bind the calyx despite conformational flexibility, but binds the dimer interface and an alternate Site C. iii) Results point to a probable lactolation site in the BLG dimer interface, at K141, consistent with previous biochemical findings. In contrast, no accessible lysines are found near Site C. iv) lactose forms hydrogen bonds with residues from both monomers stabilizing the dimer through a claw-like structure. Overall, these results improve our understanding of BLG's binding sites, importantly narrowing down the calyx residues that control ligand binding. Moreover, our results emphasize the importance of the dimer interface as an insufficiently explored, biologically relevant binding site of particular importance for hydrophilic ligands. Furthermore our analyses suggest that BLG is a robust scaffold for multiple ligand-binding, suitable for protein design, and advance our molecular understanding of its ligand sites to a point that allows manipulation to control binding.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
