

Behavioural sensitivity to binaural spatial cues in ferrets: evidence for plasticity in the duplex theory of sound localization
- PMID: 24256073
- PMCID: PMC4063341
- DOI: 10.1111/ejn.12402
Behavioural sensitivity to binaural spatial cues in ferrets: evidence for plasticity in the duplex theory of sound localization
Abstract
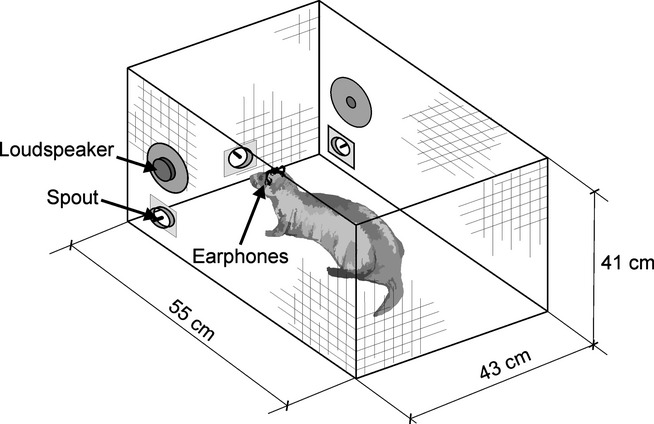
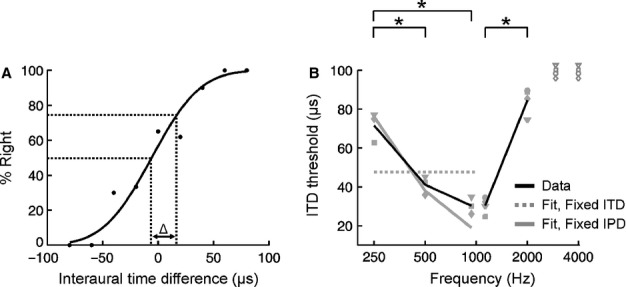
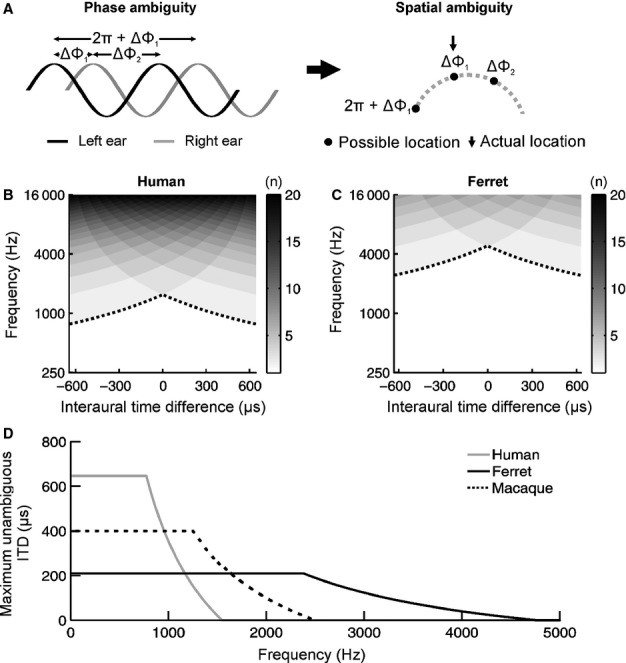
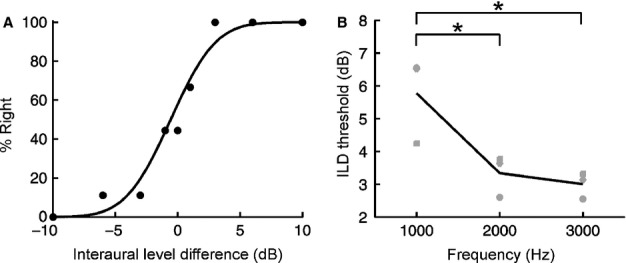
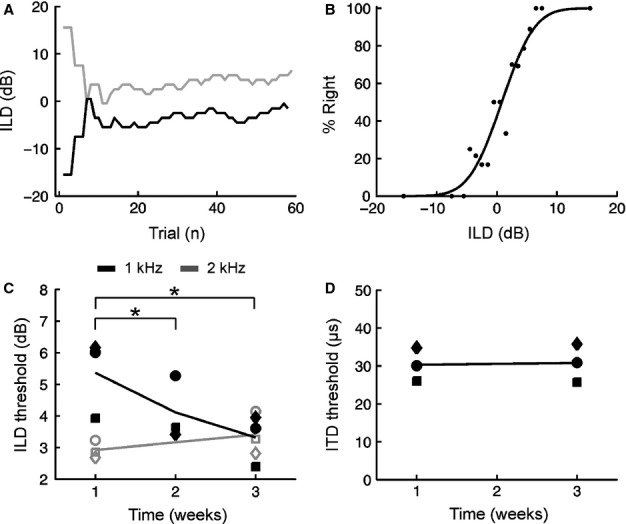
For over a century, the duplex theory has guided our understanding of human sound localization in the horizontal plane. According to this theory, the auditory system uses interaural time differences (ITDs) and interaural level differences (ILDs) to localize low-frequency and high-frequency sounds, respectively. Whilst this theory successfully accounts for the localization of tones by humans, some species show very different behaviour. Ferrets are widely used for studying both clinical and fundamental aspects of spatial hearing, but it is not known whether the duplex theory applies to this species or, if so, to what extent the frequency range over which each binaural cue is used depends on acoustical or neurophysiological factors. To address these issues, we trained ferrets to lateralize tones presented over earphones and found that the frequency dependence of ITD and ILD sensitivity broadly paralleled that observed in humans. Compared with humans, however, the transition between ITD and ILD sensitivity was shifted toward higher frequencies. We found that the frequency dependence of ITD sensitivity in ferrets can partially be accounted for by acoustical factors, although neurophysiological mechanisms are also likely to be involved. Moreover, we show that binaural cue sensitivity can be shaped by experience, as training ferrets on a 1-kHz ILD task resulted in significant improvements in thresholds that were specific to the trained cue and frequency. Our results provide new insights into the factors limiting the use of different sound localization cues and highlight the importance of sensory experience in shaping the underlying neural mechanisms.
Keywords: auditory localization; interaural level difference; interaural time difference; phase ambiguity; spatial hearing; training.
© 2013 The Authors. European Journal of Neuroscience published by Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Figures
References
-
- Algazi VR, Duda RO, Thompson DM, Avendano C. New Paltz, NY: Mohonk Mountain House; 2001. pp. 99–102. The CIPIC HRTF Database In: Proceedings of the 2001 IEEE Worskhop on Applications of Signal Processing to Audio and Acoustics.
-
- Blauert J. Spatial Hearing: The Psychophysics of Human Sound Localization. Cambridge, MA: MIT Press; 1997.
-
- Brown CH, Beecher MD, Moody DB, Stebbins WC. Localization of pure tones by old world monkeys. J. Acoust. Soc. Am. 1978;63:1484–1492. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
