

Seeing the invisible by paramagnetic and diamagnetic NMR
- PMID: 24256222
- PMCID: PMC4155411
- DOI: 10.1042/BST20130232
Seeing the invisible by paramagnetic and diamagnetic NMR
Abstract
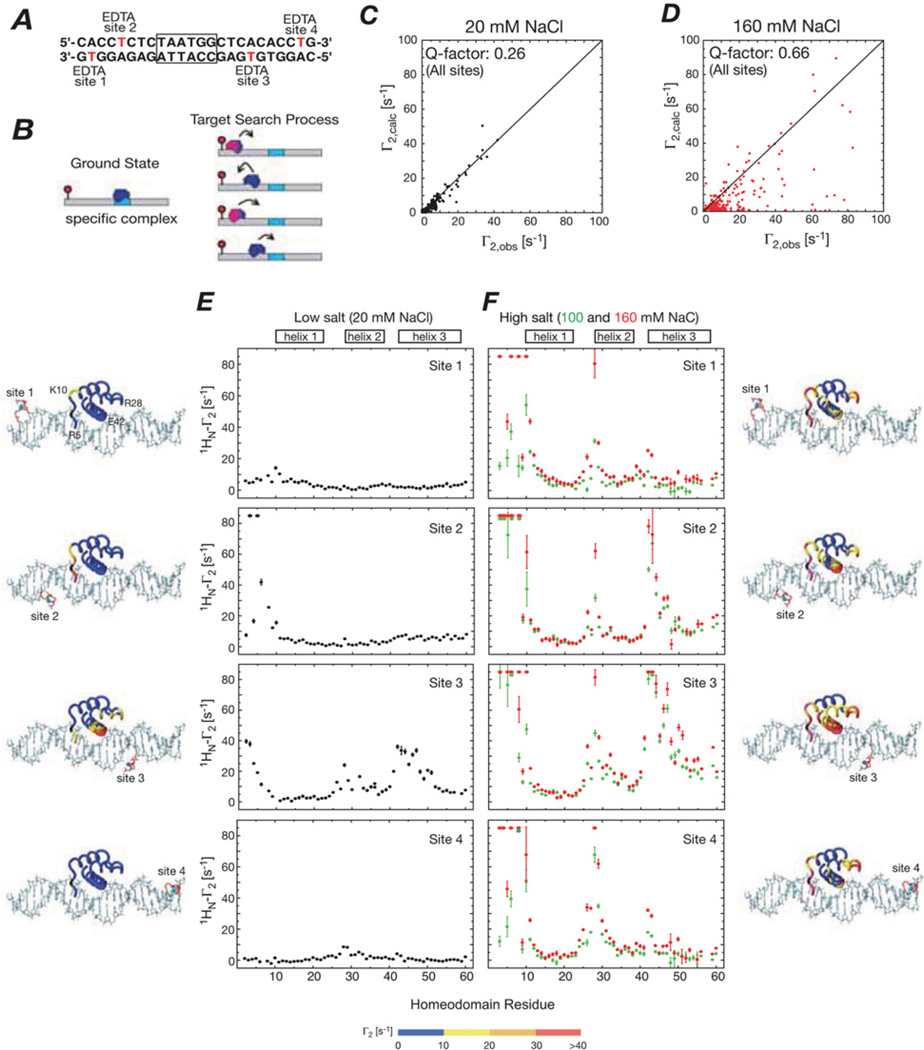
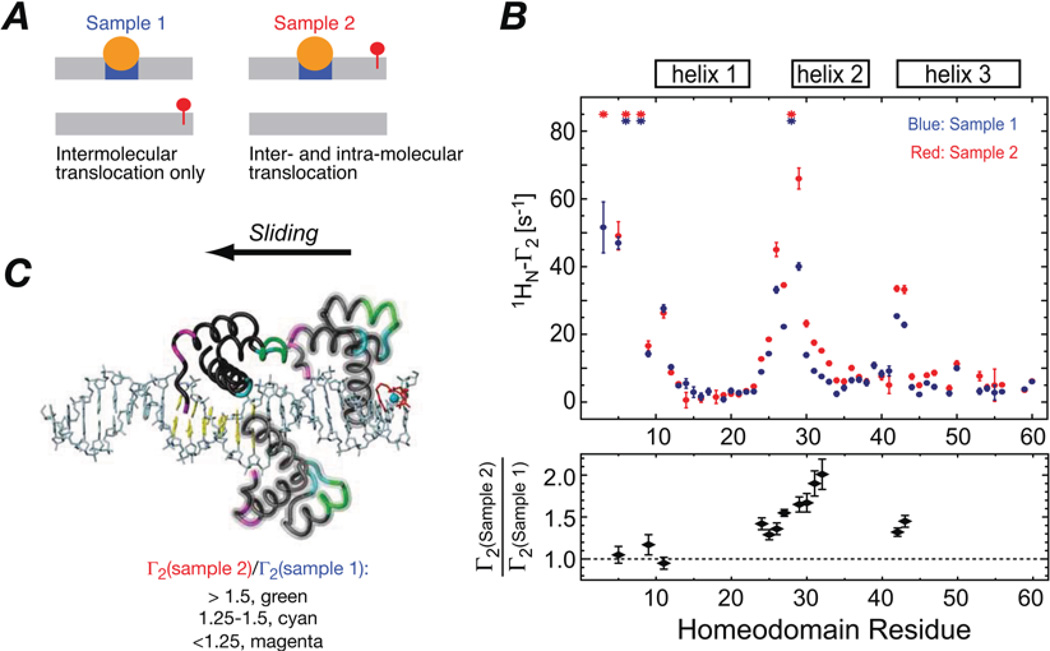
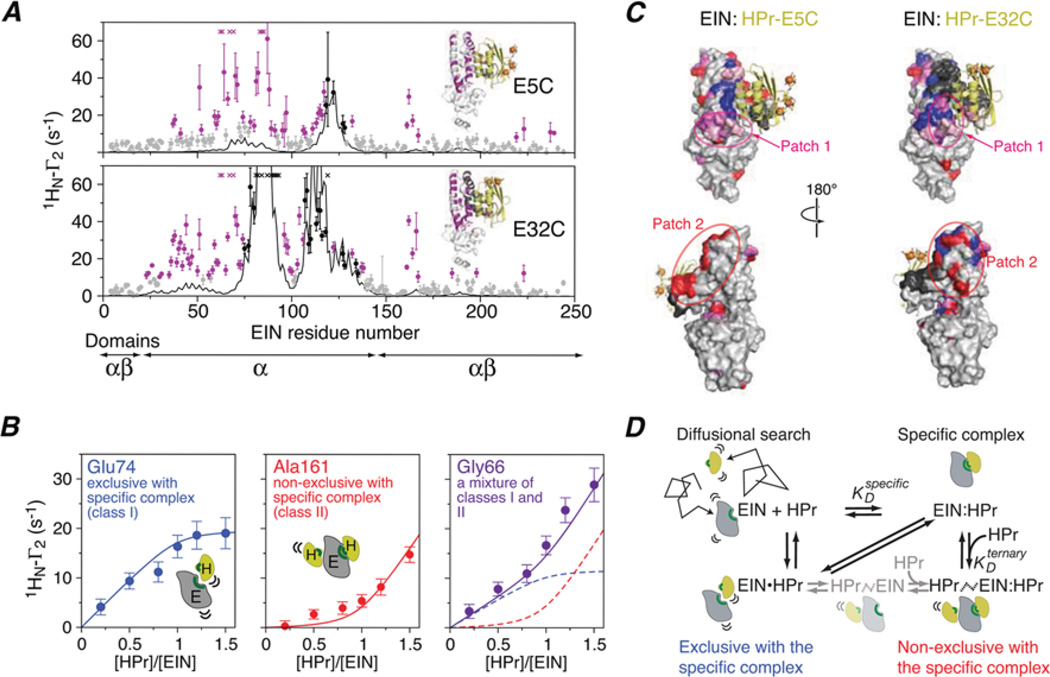
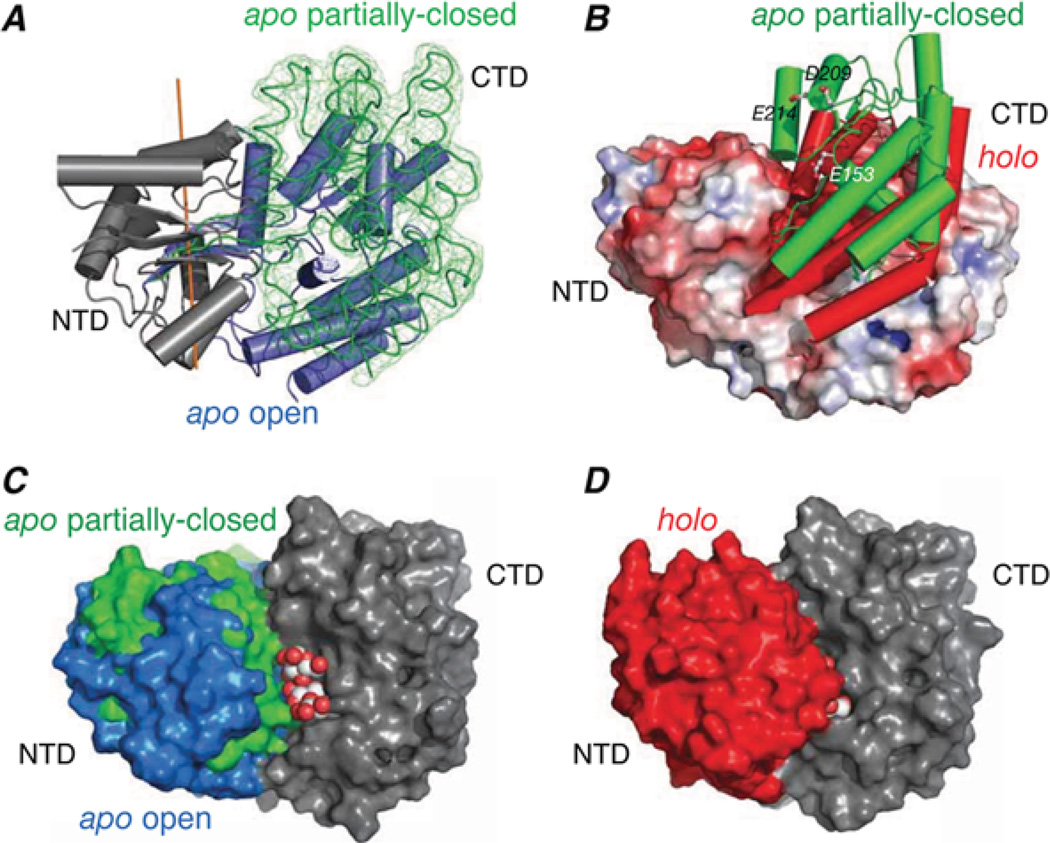
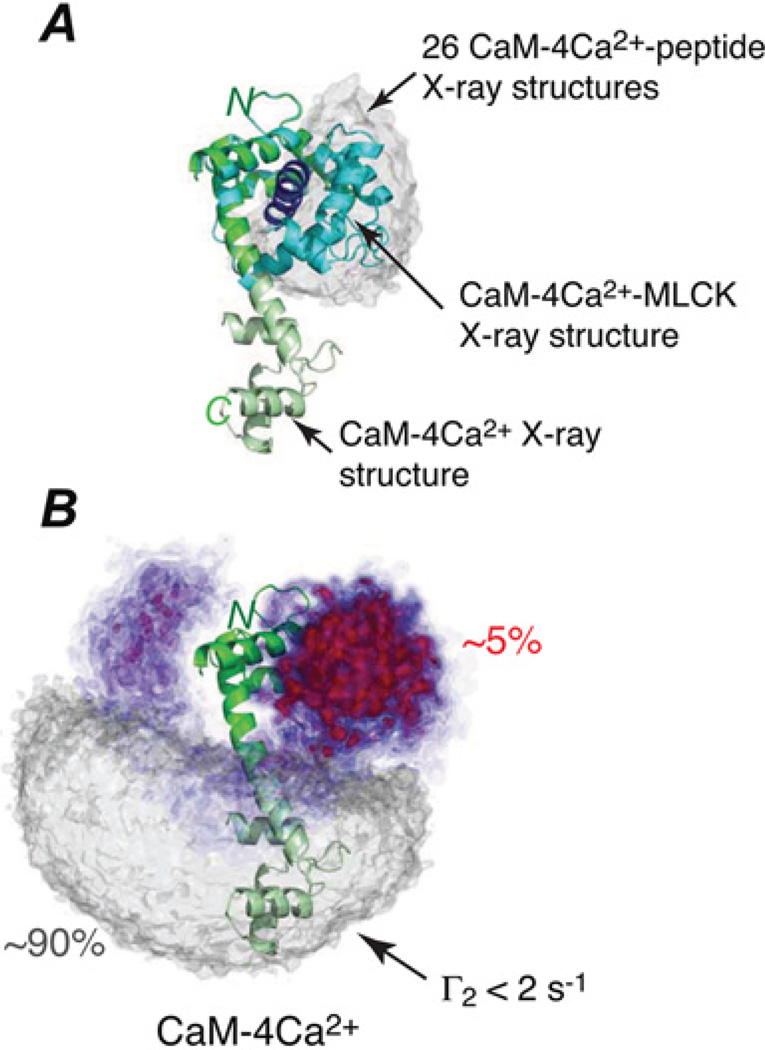
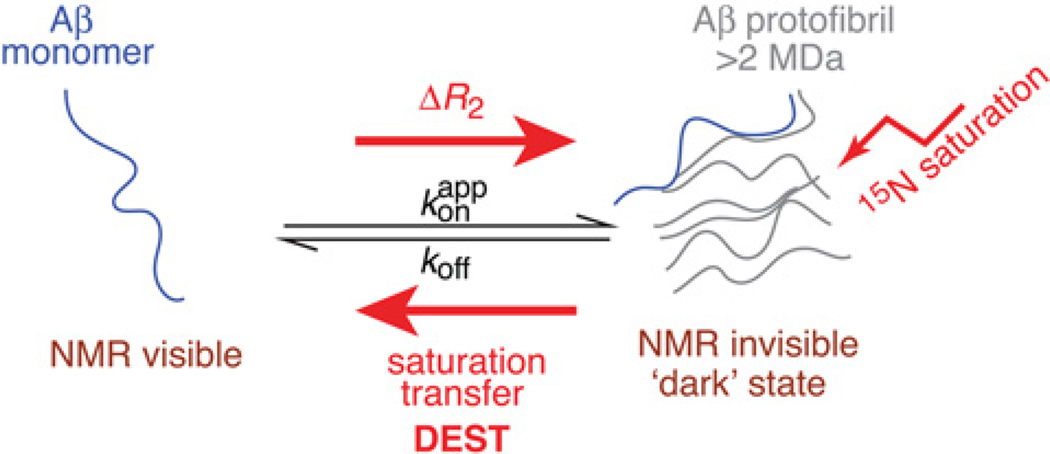
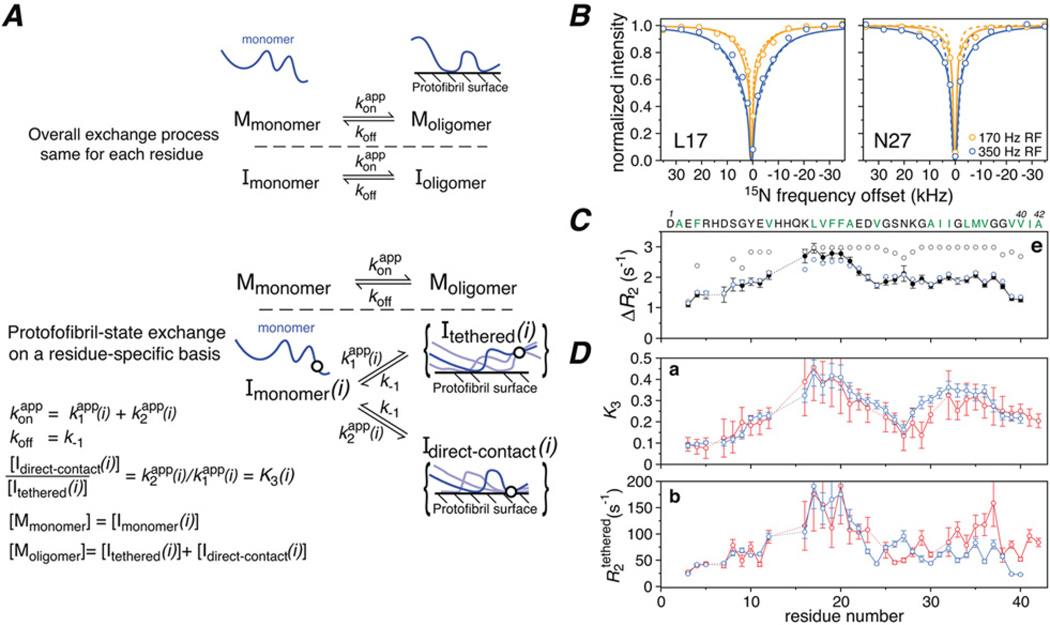
Sparsely populated transient states of proteins and their complexes play an important role in many biological processes including protein-protein and protein-DNA recognition, allostery, conformational selection, induced fit and self-assembly. These states are difficult to study as their low population and transient nature makes them effectively invisible to conventional structural and biophysical techniques. In the present article, I summarize recent NMR developments in our laboratory, including the use of paramagnetic relaxation enhancement, lifetime line broadening and dark-state exchange saturation transfer spectroscopy, that have permitted such sparsely populated states to be detected, characterized and, in some instances, visualized. I illustrate the application of these methods to the elucidation of mechanisms whereby transcription factors locate their specific target sites within an overwhelming sea of non-specific DNA, to the characterization of encounter complexes in protein-protein recognition, to large-scale interdomain motions involved in ligand binding, and to the interaction of monomeric amyloid β-peptide with the surface of amyloid protofibrils and the internal cavity surface of the chaperonin GroEL.
Figures
References
-
- Iwahara J, Clore GM. Detecting transient intermediates in macromolecular binding by paramagnetic NMR. Nature. 2006;440:1227–1230. - PubMed
-
- Palmer AG, 3rd, Kroenke CD, Loria JP. Nuclear magnetic resonance methods for quantifying microsecond-to-millisecond motions in biological macromolecules. Methods Enzymol. 2001;339:204–238. - PubMed
-
- Loria JP, Berlow RB, Watt ED. Characterization of enzyme motions by solution NMR relaxation dispersion. Acc. Chem. Res. 2008;41:214–221. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials