

Memory and modularity in cell-fate decision making
- PMID: 24256735
- PMCID: PMC4019345
- DOI: 10.1038/nature12804
Memory and modularity in cell-fate decision making
Abstract
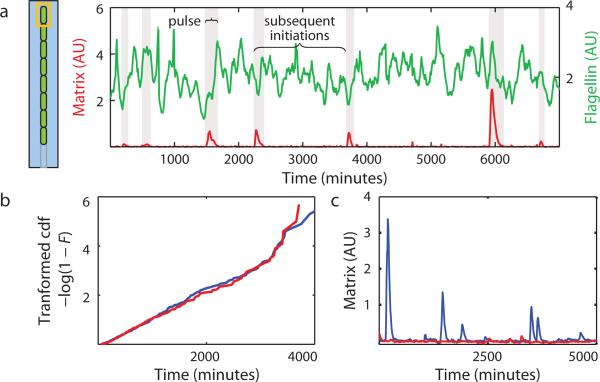
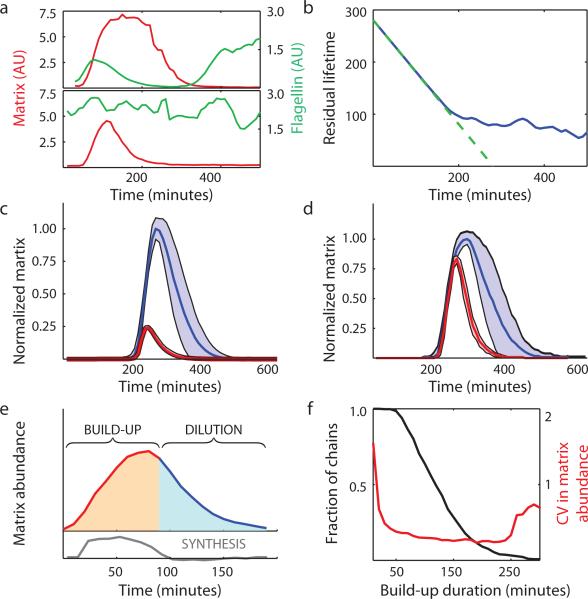
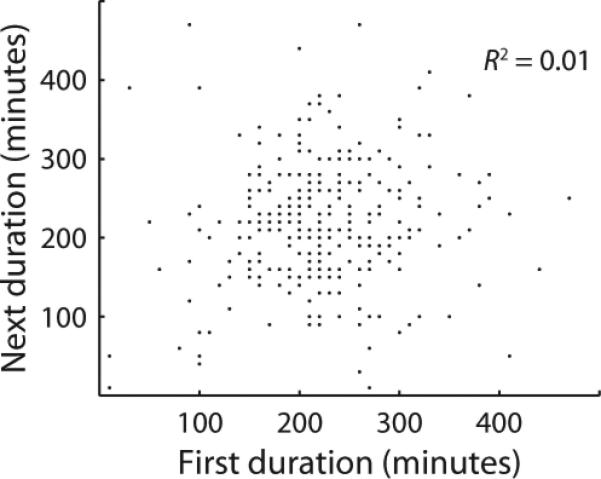
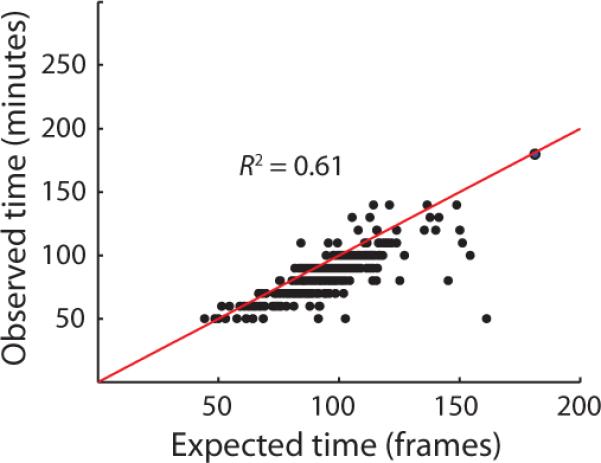
Genetically identical cells sharing an environment can display markedly different phenotypes. It is often unclear how much of this variation derives from chance, external signals, or attempts by individual cells to exert autonomous phenotypic programs. By observing thousands of cells for hundreds of consecutive generations under constant conditions, we dissect the stochastic decision between a solitary, motile state and a chained, sessile state in Bacillus subtilis. We show that the motile state is 'memoryless', exhibiting no autonomous control over the time spent in the state. In contrast, the time spent as connected chains of cells is tightly controlled, enforcing coordination among related cells in the multicellular state. We show that the three-protein regulatory circuit governing the decision is modular, as initiation and maintenance of chaining are genetically separable functions. As stimulation of the same initiating pathway triggers biofilm formation, we argue that autonomous timing allows a trial commitment to multicellularity that external signals could extend.
Figures










Comment in
-
Systems biology: How bacteria choose a lifestyle.Nature. 2013 Nov 28;503(7477):476-7. doi: 10.1038/nature12837. Epub 2013 Nov 20. Nature. 2013. PMID: 24256727 No abstract available.
References
-
- Ferrell JE, Jr, Machleder EM. The biochemical basis of an all-or-none cell fate switch in Xenopus oocytes. Science. 1998;280:895–898. - PubMed
-
- Wehrli M, Tomlinson A. Epithelial planar polarity in the developing Drosophila eye. Development. 1995;121:2451–2459. - PubMed
-
- Paulsson J. Summing up the noise in gene networks. Nature. 2004;427:415–418. - PubMed
-
- Ozbudak EM, Thattai M, Kurtser I, Grossman AD, van Oudenaarden A. Regulation of noise in the expression of a single gene. Nat. Genet. 2002;31:69–73. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
