

doi: 10.1128/JVI.03251-13.
Epub 2013 Nov 20.
A lytic viral long noncoding RNA modulates the function of a latent protein
Affiliations
- PMID: 24257619
- PMCID: PMC3911622
- DOI: 10.1128/JVI.03251-13
Item in Clipboard
A lytic viral long noncoding RNA modulates the function of a latent protein
J Virol.
2014 Feb.
Abstract
Latent Kaposi's sarcoma-associated herpesvirus (KSHV) episomes are coated with viral latency-associated nuclear antigen (LANA). In contrast, LANA rapidly disassociates from episomes during reactivation. Lytic KSHV expresses polyadenylated nuclear RNA (PAN RNA), a long noncoding RNA (lncRNA). We report that PAN RNA promotes LANA-episome disassociation through an interaction with LANA which facilitates LANA sequestration away from KSHV episomes during reactivation. These findings suggest that KSHV may have evolved an RNA aptamer to regulate latent protein function.
Figures
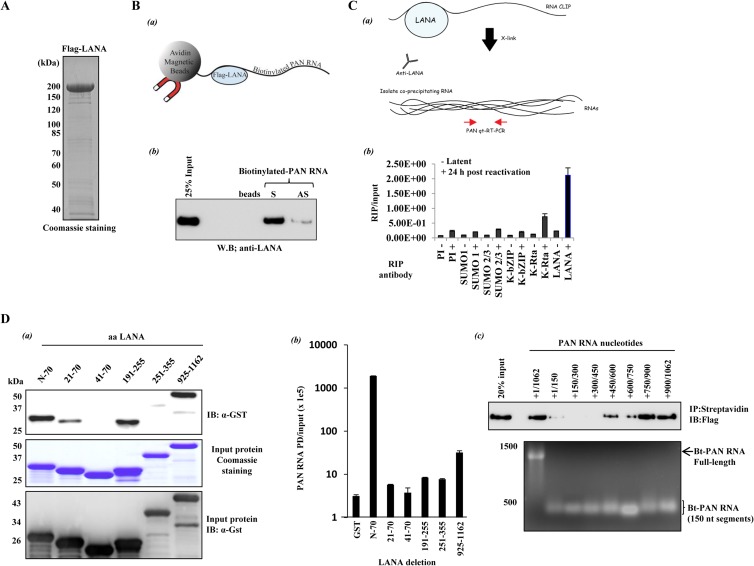
LANA-PAN RNA interaction. (A) Flag-LANA expression. Flag-LANA was produced from baculovirus-infected Sf9 cells. LANA was purified on FLAG-M2 resin and analyzed by SDS-PAGE with Coomassie brilliant blue staining. (B) In vitro LANA-PAN RNA interaction. (a) Illustration of the reaction. (b) Purified Flag-LANA was incubated with biotinylated S or AS PAN RNA. The interaction was captured with streptavidin magnetic beads and probed with anti-Flag. “beads” indicates the incubation of streptavidin beads plus Flag-LANA. (C) In vivo LANA-PAN interaction analyzed by RNA CLIP. (a) Illustration of the reaction. (b) Before (−) and after (+) 24 h chemical reactivation, BCBL-1 cells were fixed with 0.3% formaldehyde, and RNA-LANA complexes were precipitated with the indicated antibodies. After the reversal of cross-links and purification of RNA from the protein-RNA complexes, the amount of precipitated PAN RNA was measured by RT-qPCR using PAN RNA-specific primers. Values are the means and standard deviations (SD) from 3 determinations. (D) Mapping the LANA- and PAN RNA-interacting domains. (a) LANA-interacting domain. GST pulldown (PD)-protein capture. GST-LANA deletion fusion proteins were produced in Escherichia coli, purified on glutathione Sepharose, and eluted from the beads with glutathione. The fusion proteins were incubated with in vitro-transcribed biotinylated PAN RNA, and complexes were isolated with magnetic streptavidin beads. Captured protein was detected with anti-GST antibody (top). Purified proteins used in these reactions are shown by staining with Coomassie (middle) and immunoblotting with anti-GST (bottom). (b) GST-PD-RNA capture. After PD and washing, the GST fusion beads were processed for quantification of bound RNA. Captured PAN RNA was quantified by RT-qPCR. LANA amino acid residues in each deletion are listed in panels a and b. (c) Mapping of PAN RNA nucleotides that interact with LANA. PD was carried out with full-length Flag-LANA plus in vitro-transcribed, biotinylated PAN RNA fragments. The endpoints of each PAN RNA are given. Anti-Flag analysis of the pull-down (top) and denaturing agarose gel-ethidium bromide staining of the in vitro transcribed RNAs utilized (bottom) are shown.
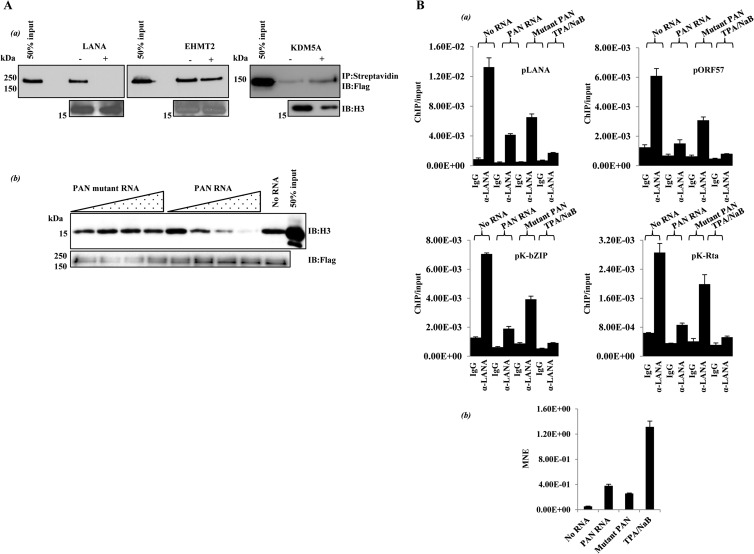
PAN RNA influences LANA-histone interaction in vitro and in vivo. (A) In vitro. (a) Histone binding reactions (14) were carried out using the purified proteins indicated. Biotinylated histone H3 (Active Motif) was used as the binding substrate. − and +, presence and absence of unlabeled PAN RNA competitor. Complexes were captured with streptavidin beads and analyzed by immunoblotting. (b) Mutant PAN lacks activity. Histone binding reaction mixtures with Flag LANA beads and histone H3 contained 0, 0.5, 5, 50, or 500 nM in vitro-transcribed, unlabeled PAN RNA or a non-LANA-interacting PAN RNA fragment (PAN nucleotides 300 to 450). After washing, bound H3 was analyzed by immunoblotting. (B) In vivo. (a) Latent 293T/rKSHV.219 cells were nucleofected (Lonza Nucleofector) with in vitro-transcribed PAN RNA or mutant PAN RNA (PAN nucleotides 300 to 450) (0.5 μM RNA), and an ORF57 expression vector DNA. Eighteen hours later, the cells were fixed as described in the Fig. 1 legend and processed for LANA ChIP, and eluted DNA was analyzed by qPCR. The amount of immunoprecipitated, amplified DNA relative to input is listed for 4 KSHV genomic loci (means and SD from 3 determinations). Chemical reactivation (3 mM sodium butyrate [NaB] and 20 ng/ml 12-O-tetradecanoylphorbol acetate [TPA]) was used as a positive control. (b) Nucleofected PAN RNA expression. Prior to fixation for ChIP, a small amount of each cell culture was harvested for RNA isolation. cDNA was prepared and analyzed for PAN or mutant PAN RNA expression by qPCR (MNE, mean normalized expression; reference, β-actin).
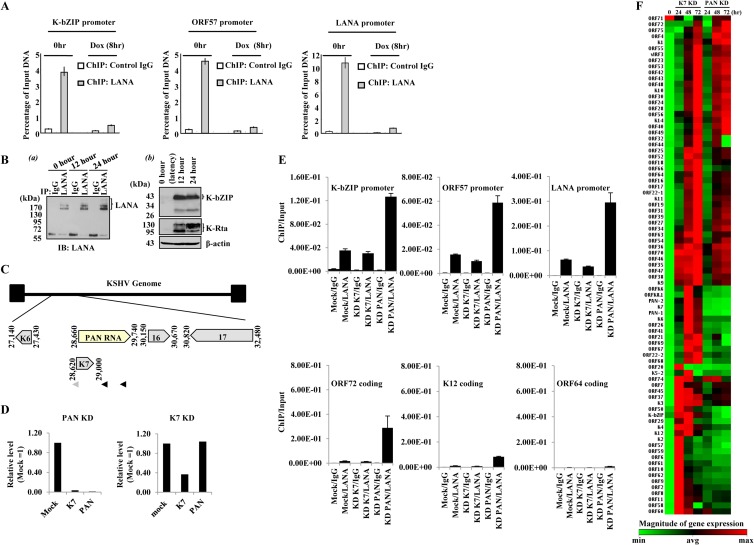
LANA-episome dynamics. (A) KSHV episome-LANA dynamics during reactivation. Anti-LANA and control IgG ChIP DNAs were prepared from K-Rta-inducible BCBL-1 cells at 0 and 8 h after K-Rta-mediated reactivation. The immunoprecipitated DNA was analyzed by qPCR using primer pairs against the KSHV ORFs listed. Values are the percentage of input DNA. Values are derived from the ratio of the threshold cycle (CT) values of the amplified DNA immunoprecipitated with each antibody relative to the amount of input DNA amplified. (B) (a) LANA protein levels during the early stages of reactivation. K-Rta-inducible BCBL-1 cell lysates were prepared at the indicated times, immunoprecipitated with the indicated antibodies, and probed with anti-LANA antibody. (b) Verification of reactivation. K-Rta-inducible BCBL-1 cell lysates were prepared at the indicated times. Immunoblots were probed with the indicated antibodies. (C) KSHV PAN RNA genomic organization. An expanded view of the K7/PAN locus is shown, and the positions of K7 KD (gray arrowhead) and PAN RNA KD oligonucleotides (black arrowheads) are shown. (D to F) PAN RNA KD alters LANA occupancy of KSHV episomes and viral gene expression. (D) PAN RNA and K7 knockdown. iSLK/rKSHV.219 cells were nucleofected with PAN or K7 antisense oligonucleotides and reactivated by doxycycline addition. At 48 h postreactivation, PAN and K7 gene expression was analyzed by qPCR. PAN RNA KD affects both K7 and PAN RNA transcript levels; K7 knockdown affects only K7 mRNAs. The expression level relative to that obtained with mock nucleofection is shown. (E) LANA ChIP. BCBL-1 cells were nucleofected with antisense PAN RNA or K7 control oligonucleotides 24 h prior to chemical reactivation. Cells were fixed with 0.3% formaldehyde after 24 h chemical reactivation, and chromatin was immunoprecipitated with control IgG or rat anti-LANA IgG. The immunoprecipitated DNA was isolated and quantitated by qPCR. The amount of immunoprecipitated DNA relative to input is listed for each KSHV locus examined. Values are the means and SD from 3 determinations. (F) KSHV gene expression array. After PAN/K7 KD and reactivation, RNA was harvested at 24, 48, and 72 h and reverse transcribed, and cDNAs were analyzed with a KSHV gene expression array. A clustergram was generated from the data set using nonsupervised hierarchical clustering. A heat map depicting relative expression levels with dendrograms is shown. RNA prepared from mock nucleofected, nonreactivated BCBL-1 cells served as the latent (0-h) control (normalized expression; green, low; red, high).
References
-
- Kapranov P, Cheng J, Dike S, Nix DA, Duttagupta R, Willingham AT, Stadler PF, Hertel J, Hackermuller J, Hofacker IL, Bell I, Cheung E, Drenkow J, Dumais E, Patel S, Helt G, Ganesh M, Ghosh S, Piccolboni A, Sementchenko V, Tammana H, Gingeras TR. 2007. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 316:1484–1488. 10.1126/science.1138341 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
