

Cell-intrinsic in vivo requirement for the E47-p21 pathway in long-term hematopoietic stem cells
- PMID: 24259504
- PMCID: PMC3893818
- DOI: 10.4049/jimmunol.1302502
Cell-intrinsic in vivo requirement for the E47-p21 pathway in long-term hematopoietic stem cells
Abstract
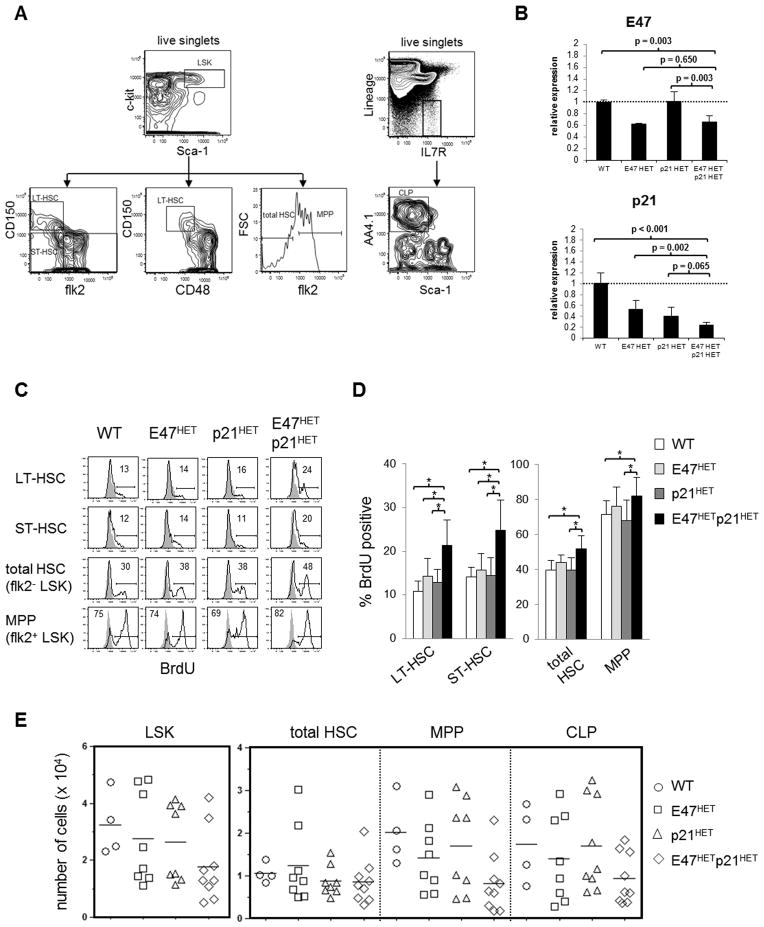
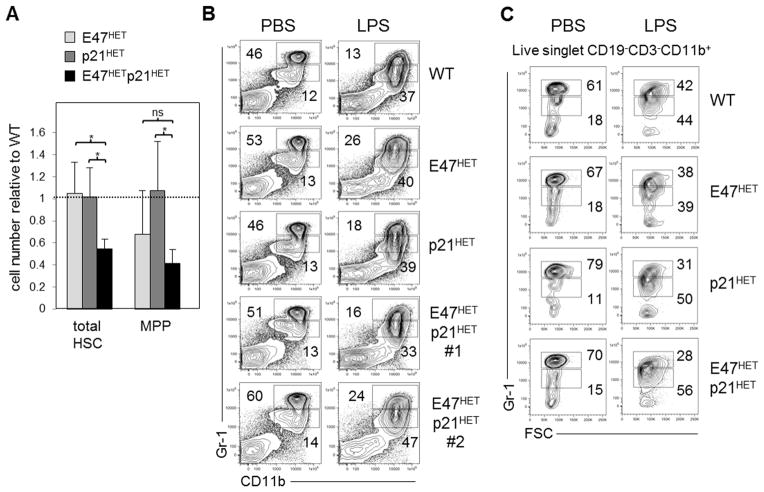
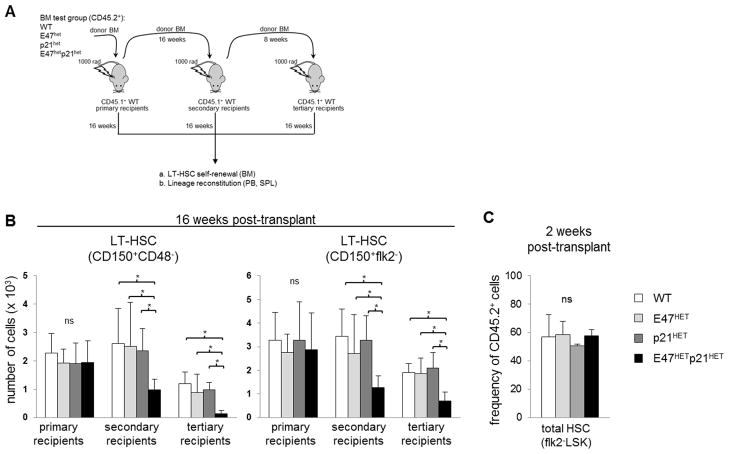
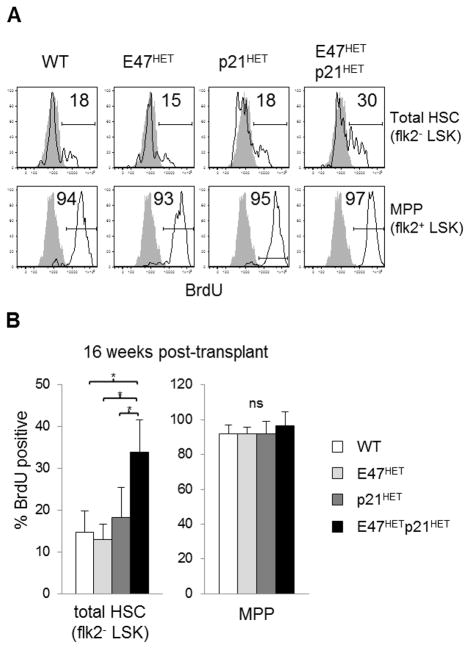
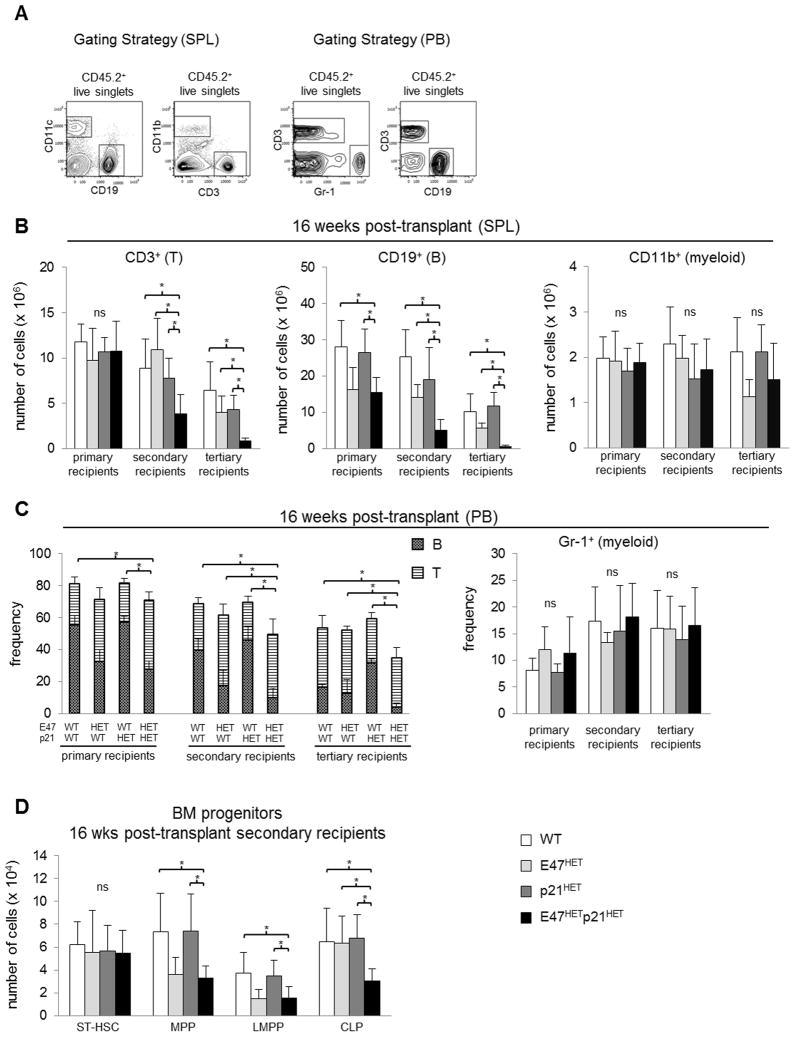
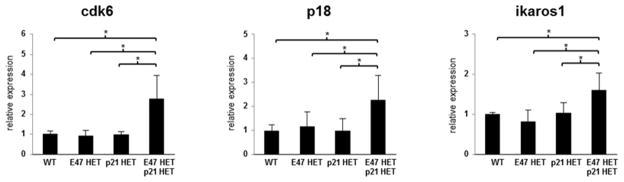
Major regulators of long-term hematopoietic stem cell (LT-HSC) self-renewal and proliferation have been identified, but knowledge of their in vivo interaction in a linear pathway is lacking. In this study, we show a direct genetic link between the transcription factor E47 and the major cell cycle regulator p21 in controlling LT-HSC integrity in vivo under repopulation stress. Numerous studies have shown that E47 activates p21 transcription in hematopoietic subsets in vitro, and we now reveal the in vivo relevance of the E47-p21 pathway by reducing the gene dose of each factor individually (E47(het) or p21(het)) versus in tandem (E47(het)p21(het)). E47(het)p21(het) LT-HSCs and downstream short-term hematopoietic stem cells exhibit hyperproliferation and preferential susceptibility to mitotoxin compared to wild-type or single haploinsufficient controls. In serial adoptive transfers that rigorously challenge self-renewal, E47(het)p21(het) LT-HSCs dramatically and progressively decline, indicating the importance of cell-intrinsic E47-p21 in preserving LT-HSCs under stress. Transient numeric recovery of downstream short-term hematopoietic stem cells enabled the production of functionally competent myeloid but not lymphoid cells, as common lymphoid progenitors were decreased, and peripheral lymphocytes were virtually ablated. Thus, we demonstrate a developmental compartment-specific and lineage-specific requirement for the E47-p21 pathway in maintaining LT-HSCs, B cells, and T cells under hematopoietic repopulation stress in vivo.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Similar articles
-
Hematopoiesis in steady-state versus stress: self-renewal, lineage fate choice, and the conversion of danger signals into cytokine signals in hematopoietic stem cells.J Immunol. 2014 Sep 1;193(5):2053-8. doi: 10.4049/jimmunol.1400936. J Immunol. 2014. PMID: 25128551 Free PMC article. Review.
-
E47 regulates hematopoietic stem cell proliferation and energetics but not myeloid lineage restriction.Blood. 2011 Mar 31;117(13):3529-38. doi: 10.1182/blood-2010-07-297689. Epub 2011 Jan 27. Blood. 2011. PMID: 21273306 Free PMC article.
-
E47 controls the developmental integrity and cell cycle quiescence of multipotential hematopoietic progenitors.J Immunol. 2008 Nov 1;181(9):5885-94. doi: 10.4049/jimmunol.181.9.5885. J Immunol. 2008. PMID: 18941177 Free PMC article.
-
Reduced lymphoid lineage priming promotes human hematopoietic stem cell expansion.Cell Stem Cell. 2014 Jan 2;14(1):94-106. doi: 10.1016/j.stem.2013.11.021. Cell Stem Cell. 2014. PMID: 24388174
-
Transcriptional and epigenetic regulation of B cell development.Immunol Res. 2011 Aug;50(2-3):105-12. doi: 10.1007/s12026-011-8225-y. Immunol Res. 2011. PMID: 21717070 Review.
Cited by
-
Bone Marrow-Derived Mesenchymal Stromal Cells from Patients with Sickle Cell Disease Display Intact Functionality.Biol Blood Marrow Transplant. 2017 May;23(5):736-745. doi: 10.1016/j.bbmt.2017.01.081. Epub 2017 Jan 26. Biol Blood Marrow Transplant. 2017. PMID: 28132869 Free PMC article.
-
Hematopoiesis in steady-state versus stress: self-renewal, lineage fate choice, and the conversion of danger signals into cytokine signals in hematopoietic stem cells.J Immunol. 2014 Sep 1;193(5):2053-8. doi: 10.4049/jimmunol.1400936. J Immunol. 2014. PMID: 25128551 Free PMC article. Review.
-
The Concerted Action of E2-2 and HEB Is Critical for Early Lymphoid Specification.Front Immunol. 2019 Mar 18;10:455. doi: 10.3389/fimmu.2019.00455. eCollection 2019. Front Immunol. 2019. PMID: 30936870 Free PMC article.
-
Transcription elongation factor ELL2 drives Ig secretory-specific mRNA production and the unfolded protein response.J Immunol. 2014 Nov 1;193(9):4663-74. doi: 10.4049/jimmunol.1401608. Epub 2014 Sep 19. J Immunol. 2014. PMID: 25238757 Free PMC article.
-
Sex-, Stress-, and Sympathetic Post-Ganglionic Neuron-Dependent Changes in the Expression of Pro- and Anti-Inflammatory Mediators in Rat Dural Immune Cells.Headache. 2015 Jul-Aug;55(7):943-57. doi: 10.1111/head.12596. Epub 2015 Jun 30. Headache. 2015. PMID: 26126992 Free PMC article.
References
-
- Adolfsson J, Mansson R, Buza-Vidas N, Hultquist A, Liuba K, Jensen CT, Bryder D, Yang L, Borge OJ, Thoren LA, Anderson K, Sitnicka E, Sasaki Y, Sigvardsson M, Jacobsen SE. Identification of Flt3+ lympho-myeloid stem cells lacking erythro-megakaryocytic potential a revised road map for adult blood lineage commitment. Cell. 2005;121:295–306. - PubMed
-
- Lai AY, Lin SM, Kondo M. Heterogeneity of Flt3-expressing multipotent progenitors in mouse bone marrow. J Immunol. 2005;175:5016–5023. - PubMed
-
- Kee BL. E and ID proteins branch out. Nat Rev Immunol. 2009;9:175–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
