

Gli activity is critical at multiple stages of embryonic mammary and nipple development
- PMID: 24260306
- PMCID: PMC3832531
- DOI: 10.1371/journal.pone.0079845
Gli activity is critical at multiple stages of embryonic mammary and nipple development
Abstract
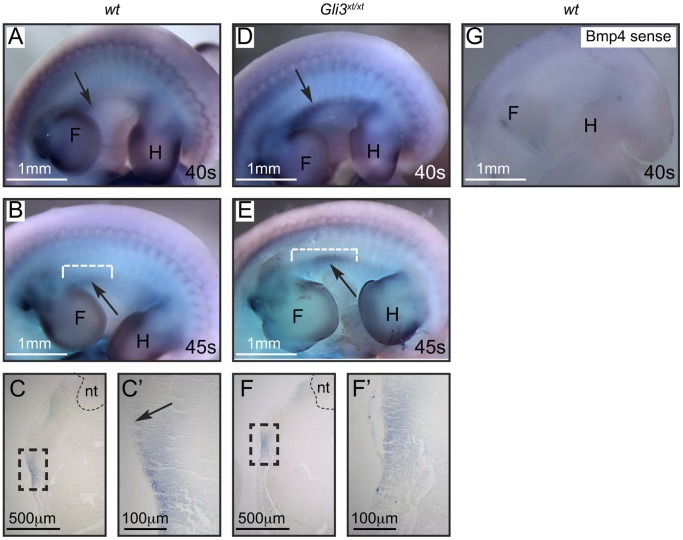
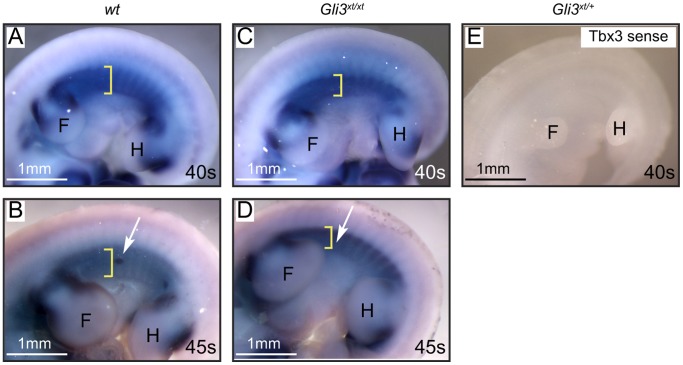
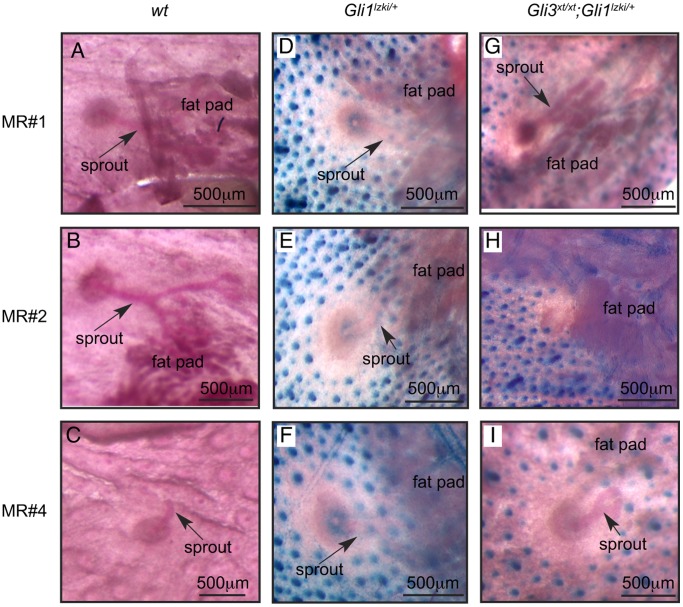
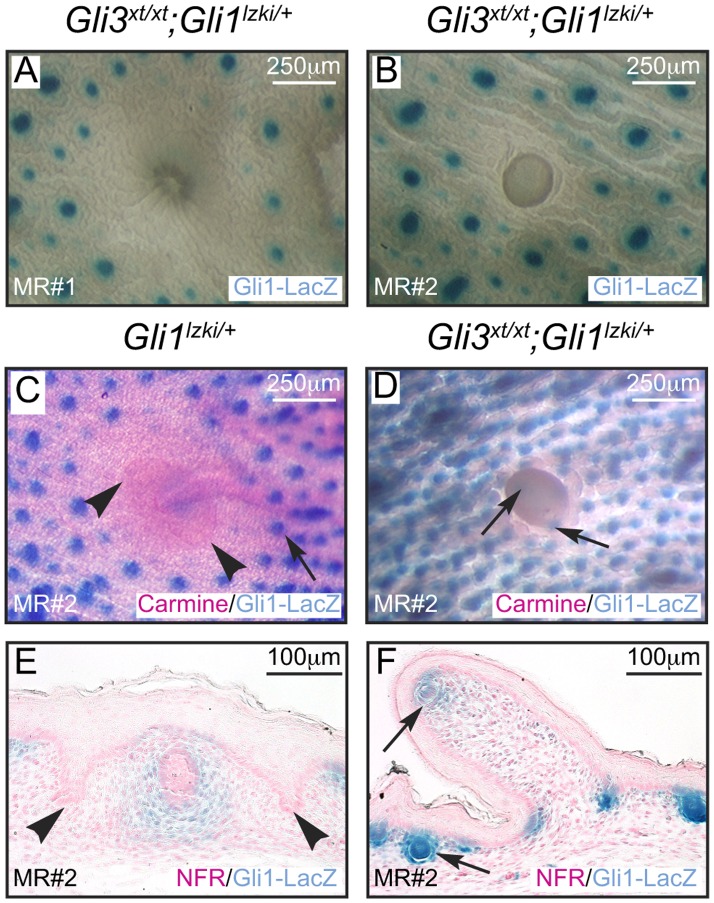
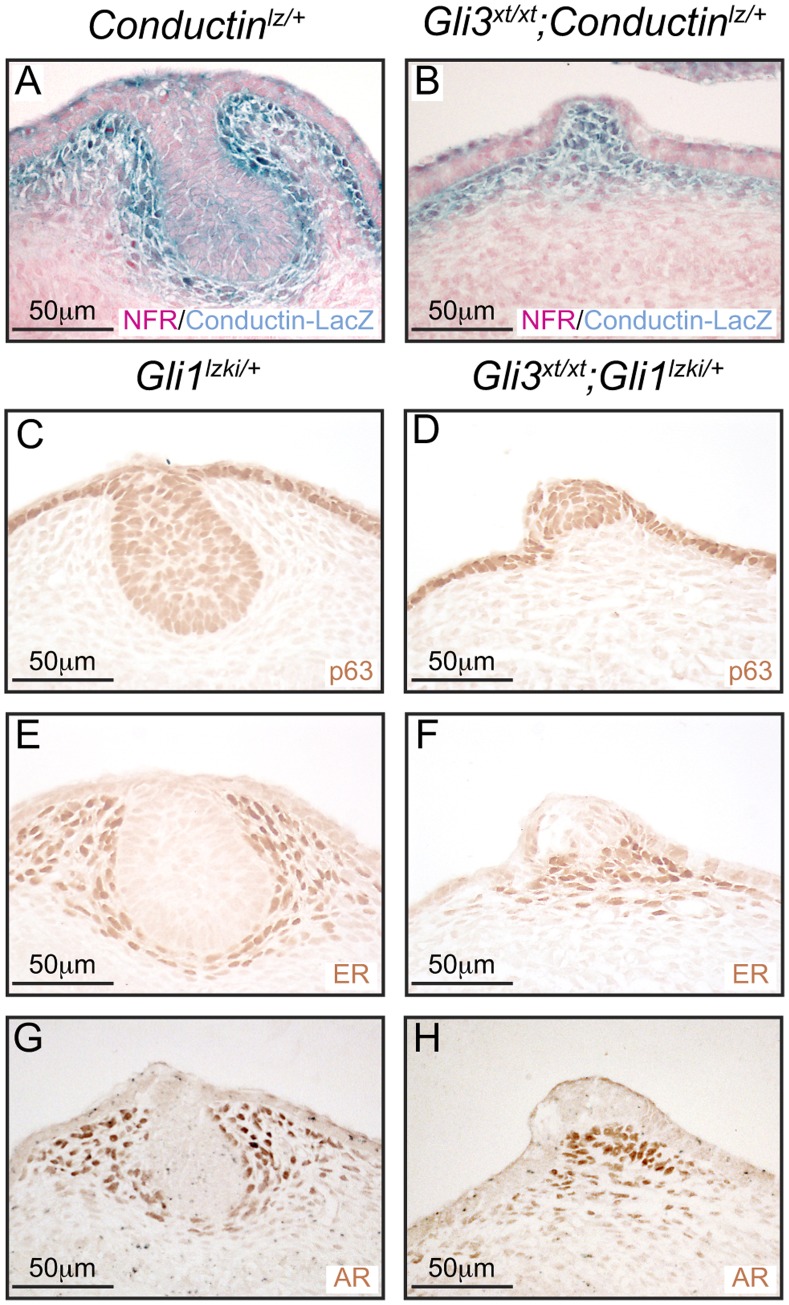
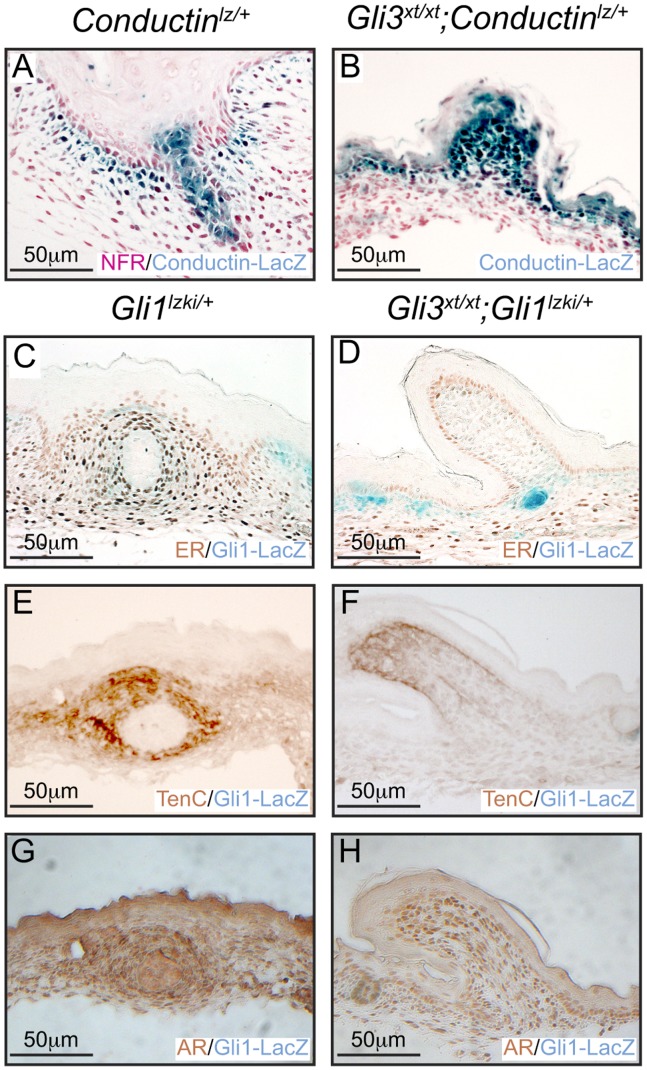
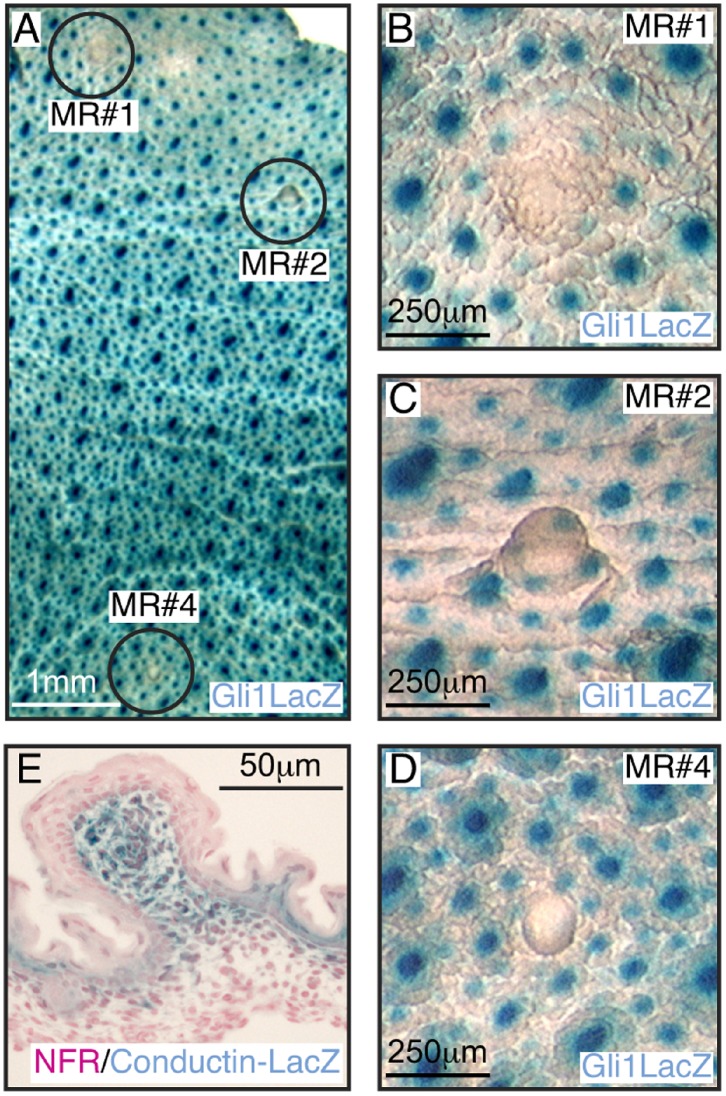
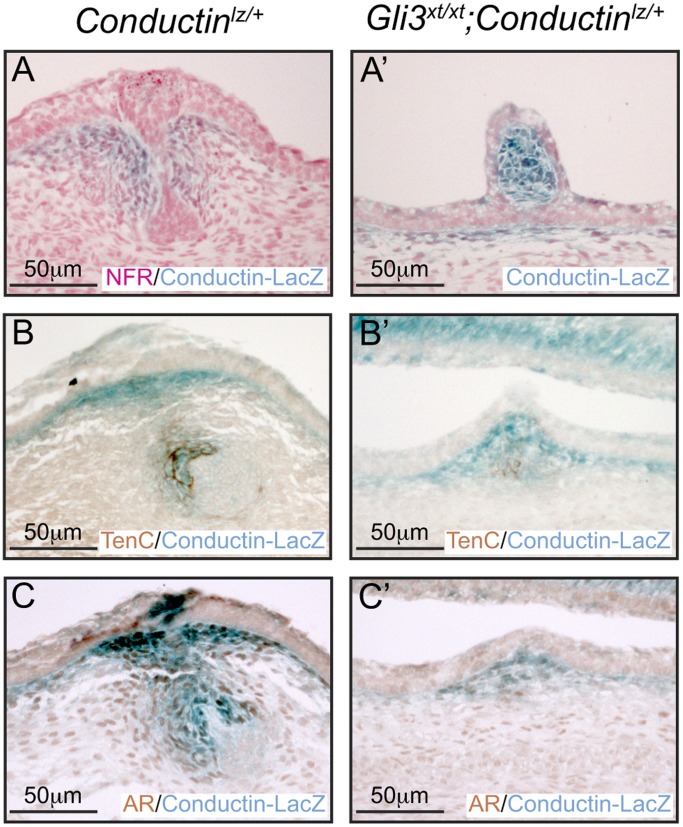
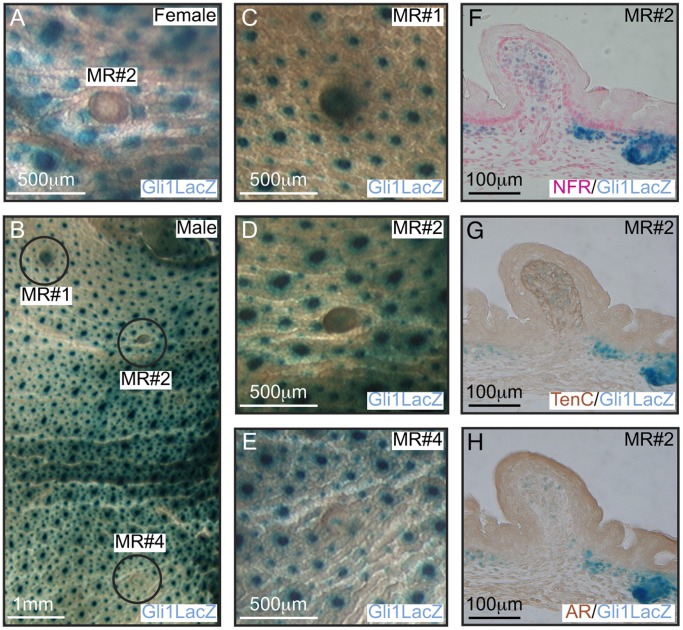
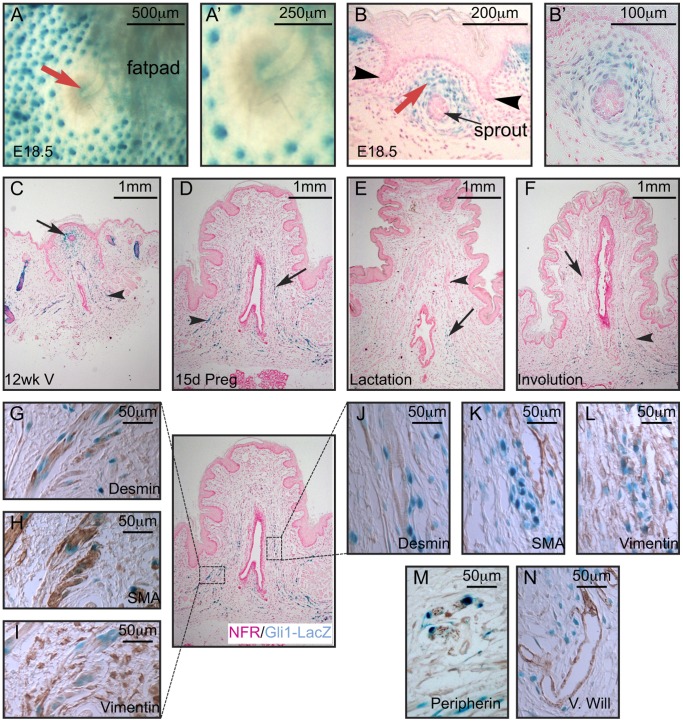
Gli3 is a transcriptional regulator of Hedgehog (Hh) signaling that functions as a repressor (Gli3(R)) or activator (Gli3(A)) depending upon cellular context. Previously, we have shown that Gli3(R) is required for the formation of mammary placodes #3 and #5. Here, we report that this early loss of Gli3 results in abnormal patterning of two critical regulators: Bmp4 and Tbx3, within the presumptive mammary rudiment (MR) #3 zone. We also show that Gli3 loss leads to failure to maintain mammary mesenchyme specification and loss of epithelial Wnt signaling, which impairs the later development of remaining MRs: MR#2 showed profound evagination and ectopic hairs formed within the presumptive areola; MR#4 showed mild invagination defects and males showed inappropriate retention of mammary buds in Gli3(xt/xt) mice. Importantly, mice genetically manipulated to misactivate Hh signaling displayed the same phenotypic spectrum demonstrating that the repressor function of Gli3(R) is essential during multiple stages of mammary development. In contrast, positive Hh signaling occurs during nipple development in a mesenchymal cuff around the lactiferous duct and in muscle cells of the nipple sphincter. Collectively, these data show that repression of Hh signaling by Gli3(R) is critical for early placodal patterning and later mammary mesenchyme specification whereas positive Hh signaling occurs during nipple development.
Conflict of interest statement
Figures
Similar articles
-
Gli3-mediated repression of Hedgehog targets is required for normal mammary development.Development. 2006 Sep;133(18):3661-70. doi: 10.1242/dev.02542. Epub 2006 Aug 16. Development. 2006. PMID: 16914490
-
BMP4 and PTHrP interact to stimulate ductal outgrowth during embryonic mammary development and to inhibit hair follicle induction.Development. 2007 Mar;134(6):1221-30. doi: 10.1242/dev.000182. Epub 2007 Feb 14. Development. 2007. PMID: 17301089
-
GLI3-dependent transcriptional repression of Gli1, Gli2 and kidney patterning genes disrupts renal morphogenesis.Development. 2006 Feb;133(3):569-78. doi: 10.1242/dev.02220. Epub 2006 Jan 5. Development. 2006. PMID: 16396903
-
Hedgehog and Gli signaling in embryonic mammary gland development.J Mammary Gland Biol Neoplasia. 2013 Jun;18(2):133-8. doi: 10.1007/s10911-013-9291-7. Epub 2013 May 16. J Mammary Gland Biol Neoplasia. 2013. PMID: 23677624 Free PMC article. Review.
-
Hedgehog-GLI mediated control of renal formation and malformation.Front Nephrol. 2023 Apr 20;3:1176347. doi: 10.3389/fneph.2023.1176347. eCollection 2023. Front Nephrol. 2023. PMID: 37675356 Free PMC article. Review.
Cited by
-
Hoxc8 initiates an ectopic mammary program by regulating Fgf10 and Tbx3 expression and Wnt/β-catenin signaling.Development. 2015 Dec 1;142(23):4056-67. doi: 10.1242/dev.128298. Epub 2015 Oct 12. Development. 2015. PMID: 26459221 Free PMC article.
-
Fetal Mammary Gland Development and Offspring's Breast Cancer Risk in Adulthood.Biology (Basel). 2025 Jan 21;14(2):106. doi: 10.3390/biology14020106. Biology (Basel). 2025. PMID: 40001874 Free PMC article. Review.
-
The Hedgehog Signaling Pathway: A Viable Target in Breast Cancer?Cancers (Basel). 2019 Aug 7;11(8):1126. doi: 10.3390/cancers11081126. Cancers (Basel). 2019. PMID: 31394751 Free PMC article. Review.
-
The Stromal Niche for Epithelial Stem Cells: A Template for Regeneration and a Brake on Malignancy.Cancer Cell. 2017 Oct 9;32(4):404-410. doi: 10.1016/j.ccell.2017.08.007. Cancer Cell. 2017. PMID: 29017054 Free PMC article. Review.
-
Normal mammary development and function in mice with Ift88 deleted in MMTV- and K14-Cre expressing cells.Cilia. 2014 Mar 4;3(1):4. doi: 10.1186/2046-2530-3-4. Cilia. 2014. PMID: 24594320 Free PMC article.
References
-
- Veltmaat JM, Mailleux AA, Thiery JP, Bellusci S (2003) Mouse embryonic mammogenesis as a model for the molecular regulation of pattern formation. Differentiation 71: 1–17. - PubMed
-
- Veltmaat JM, Van Veelen W, Thiery JP, Bellusci S (2004) Identification of the mammary line in mouse by Wnt10b expression. Dev Dyn 229: 349–356. - PubMed
-
- van Genderen C, Okamura RM, Farinas I, Quo R-G, Parslow TG, et al. (1994) Development of several organs that require inductive epithelial-mesenchymal interactions is impaired in Lef-1 deficient mice. Genes and Development 8: 2691–2704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
