

WNT5A encodes two isoforms with distinct functions in cancers
- PMID: 24260410
- PMCID: PMC3832467
- DOI: 10.1371/journal.pone.0080526
WNT5A encodes two isoforms with distinct functions in cancers
Abstract
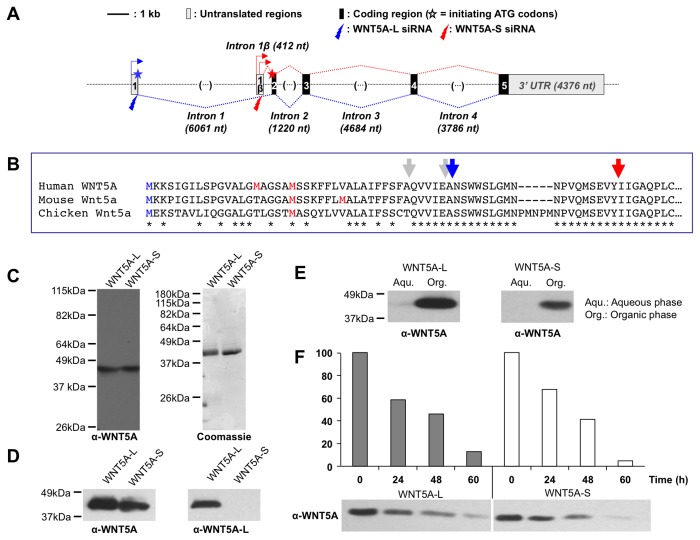
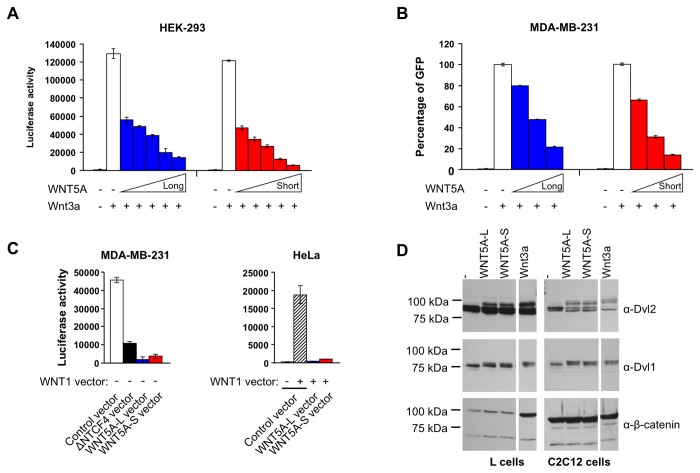
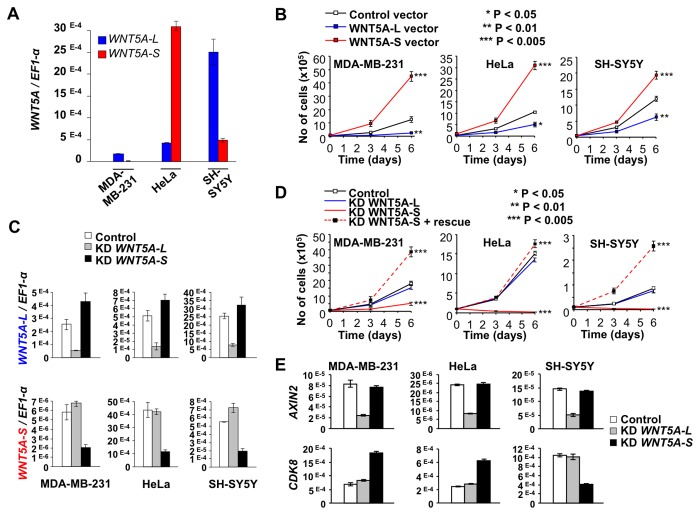
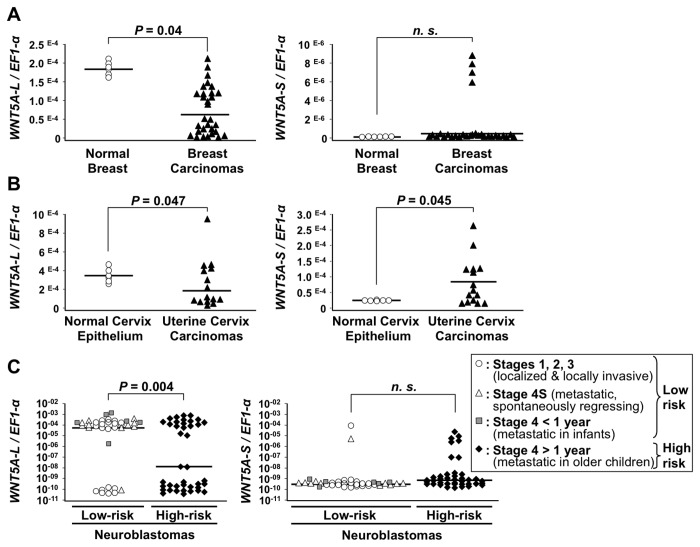
WNT5A, a member of the WNT family of secreted lipid-modified glycoproteins, is a critical regulator of a host of developmental processes, including limb formation, lung morphogenesis, intestinal elongation and mammary gland development. Altered WNT5A expression has been associated with a number of cancers. Interestingly, in certain types of cancers, such as hematological malignancies and colorectal carcinoma, WNT5A is inactivated and exerts a tumor suppressive function, while in other cancers, such as melanoma and gastric carcinoma, WNT5A is overexpressed and promotes tumor progression. The mechanism by which WNT5A achieves these distinct activities in cancers is poorly understood. Here, we provide evidence that the WNT5A gene produces two protein isoforms, WNT5A-long (WNT5A-L) and WNT5A-short (WNT5A-S). Amino-terminal sequencing and a WNT5A-L specific antibody demonstrate that the mature and secreted isoforms are distinct, with WNT5A-L carrying an additional 18 N-terminal amino acids. Biochemical analysis indicates that both purified proteins are similar with respect to their stability, hydrophobicity and WNT/β-catenin signaling activity. Nonetheless, modulation of these two WNT5A isoforms, either through ectopic expression or knockdown, demonstrates that they exert distinct activities in cancer cell lines: while WNT5A-L inhibits proliferation of tumor cell lines, WNT5A-S promotes their growth. Finally, we show that expression of these two WNT5A isoforms is altered in breast and cervix carcinomas, as well as in the most aggressive neuroblastoma tumors. In these cancers, WNT5A-L is frequently down-regulated, whereas WNT5A-S is found overexpressed in a significant fraction of tumors. Altogether, our study provides evidence that the distinct activities of WNT5A in cancer can be attributed to the production of two WNT5A isoforms.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
