

Large-scale information flow in conscious and unconscious states: an ECoG study in monkeys
- PMID: 24260491
- PMCID: PMC3829858
- DOI: 10.1371/journal.pone.0080845
Large-scale information flow in conscious and unconscious states: an ECoG study in monkeys
Erratum in
- PLoS One. 2014;9(3):e91438
Abstract
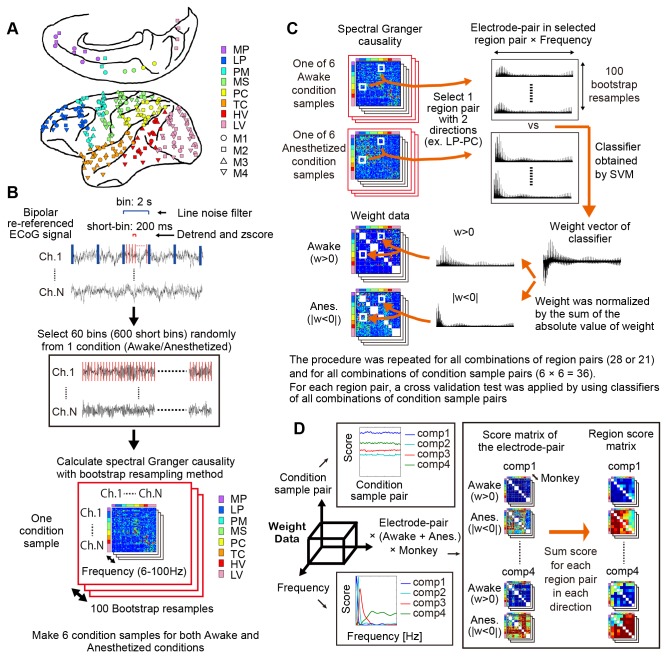
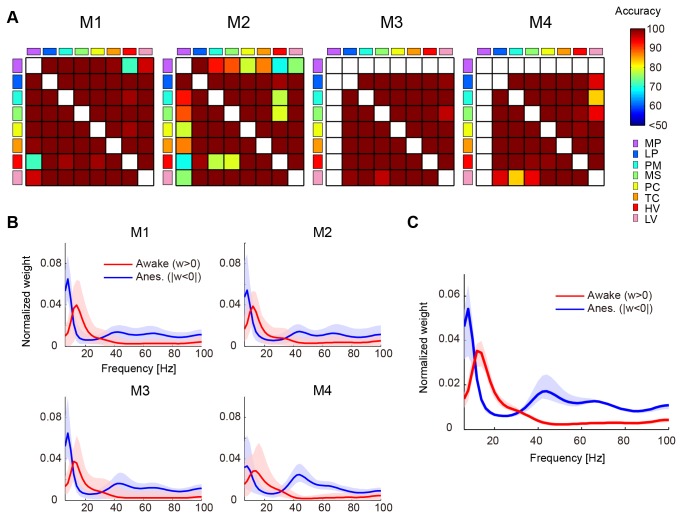
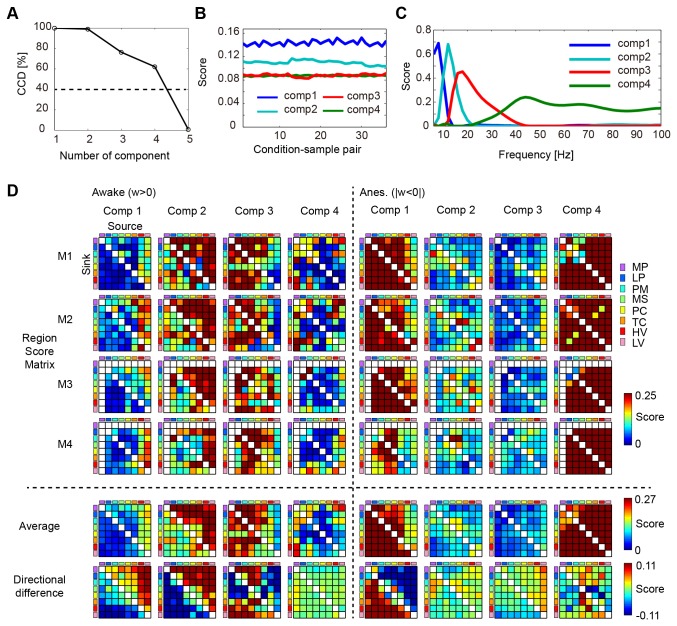
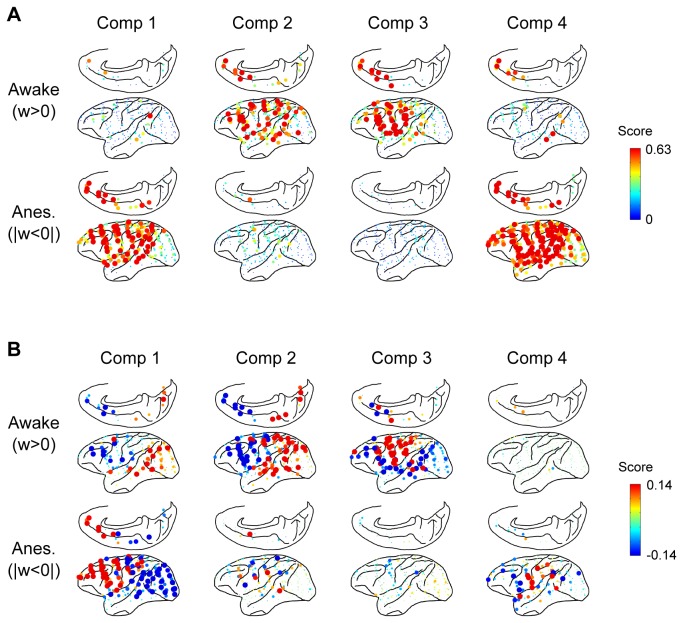
Consciousness is an emergent property of the complex brain network. In order to understand how consciousness is constructed, neural interactions within this network must be elucidated. Previous studies have shown that specific neural interactions between the thalamus and frontoparietal cortices; frontal and parietal cortices; and parietal and temporal cortices are correlated with levels of consciousness. However, due to technical limitations, the network underlying consciousness has not been investigated in terms of large-scale interactions with high temporal and spectral resolution. In this study, we recorded neural activity with dense electrocorticogram (ECoG) arrays and used the spectral Granger causality to generate a more comprehensive network that relates to consciousness in monkeys. We found that neural interactions were significantly different between conscious and unconscious states in all combinations of cortical region pairs. Furthermore, the difference in neural interactions between conscious and unconscious states could be represented in 4 frequency-specific large-scale networks with unique interaction patterns: 2 networks were related to consciousness and showed peaks in alpha and beta bands, while the other 2 networks were related to unconsciousness and showed peaks in theta and gamma bands. Moreover, networks in the unconscious state were shared amongst 3 different unconscious conditions, which were induced either by ketamine and medetomidine, propofol, or sleep. Our results provide a novel picture that the difference between conscious and unconscious states is characterized by a switch in frequency-specific modes of large-scale communications across the entire cortex, rather than the cessation of interactions between specific cortical regions.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
