

Review
doi: 10.1155/2013/752384.
eCollection 2013.
Production, control, and visual guidance of saccadic eye movements
Affiliations
- PMID: 24260720
- PMCID: PMC3821953
- DOI: 10.1155/2013/752384
Item in Clipboard
Review
Production, control, and visual guidance of saccadic eye movements
ISRN Neurol.
.
Abstract
Primate vision is served by rapid shifts of gaze called saccades. This review will survey current knowledge and particular problems concerning the neural control and guidance of gaze shifts.
Figures
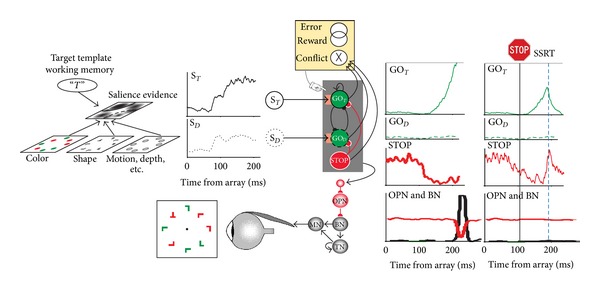
Neural networks for the guidance and control of visually guided saccades. Consider visual search for a red “T” among randomly oriented red and green “L”s. The color and shape of the objects are specified in feature maps that could also represent motion, depth, and other visual features. These feature maps converge on a map that represents the evidence for salience at each location. This salience map is also informed by a target template in working memory. The timecourse of the salience evidence representation at the target location (S
T, solid line) and a distractor location (S
D, dotted line) is plotted. According to the gated accumulator model, this evidence is integrated by a network of mutually inhibitory units that will produce a saccade to the target (GOT, solid line) or to a distractor (GOD, dotted line). A gate (orange box) prevents integration of noise by requiring the salience evidence to be of sufficient magnitude. A saccade is produced when the activation of a GO unit reaches a threshold (gray horizontal line) at which point inhibition is imposed on omnipause (OPN) neurons (red line) that releases inhibition of burst neurons (BNs) that innervate motor neurons (MNs) to produce a pulse of force to rotate the eye rapidly. The eye velocity signal from the BNs is integrated by a network of tonic neurons (TNs) that also innervate the MN to establish a step of force necessary to maintain eccentric fixation of the target. The activation of the GO units is also influenced by gaze-holding STOP units that release inhibition on the GO units while saccade preparation transpires. If a stop signal of some kind occurs, then the STOP units potently interrupt the GO unit activation from reaching the threshold; this interruption occurs within the theoretical interval known as stop signal reaction time (SSRT) (rightmost columns). An executive control network (yellow) comprised of neurons sensitive to errors, reward, and the conflict arising from coactivation of mutually incompatible response processes signals the consequences and conditions of an action. This executive control network may influence the level of the gate that systematically changes the beginning of the accumulation process to emphasize either speed or accuracy in task performance.
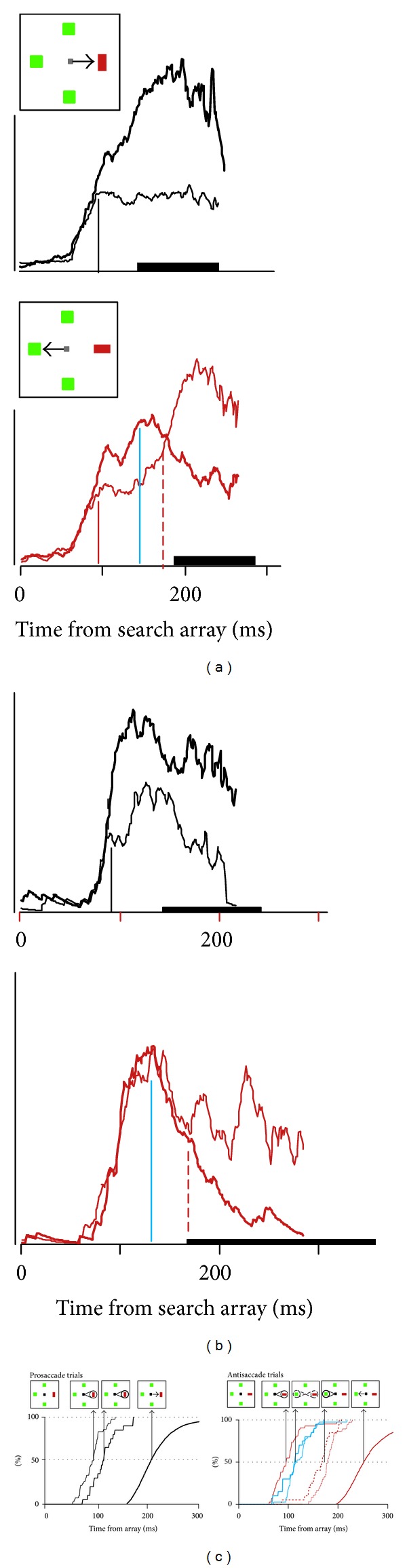
Pattern and timing of neural activity in FEF when mapping between location of visual target and endpoint of saccade is various. (a) Activity of FEF neuron with activity that can be identified with the allocation of attention (Type I). Average spike density function when the singleton fell in the neuron's receptive field (thick line) and when the singleton was located opposite the receptive field (thin line) in prosaccade (top) and antisaccade (bottom) trials. Thick bar on abscissa marks range of RT. Scale bar represents 100 spikes/sec. (b) Activity of FEF neuron with activity that can be identified with selection of the saccade endpoint (Type II). (c) Cumulative distributions of modulation times in prosaccade (left) and antisaccade (right) trials for Type I (thin) and Type II (thicker) neurons with corresponding RT (thickest). The inset arrays indicate hypothesized functional correlates. After presentation of the array, selection of the singleton location occurs first in Type I neurons (indicated by the spotlight on the singleton); this occurs at the same time in prosaccade and antisaccade trials and does not relate to whether or when gaze shifts. In prosaccade but not antisaccade trials, Type II neurons select the singleton at a later time which accounts for some of the variability of RT. A comparison of activation in prosaccade and antisaccade trials reveals the time at which the shape of the singleton is encoded to specify the correct saccade direction; this follows singleton selection and coincides for Type I (thin blue) and Type II (thicker blue) neurons in antisaccade trials. At this moment in antisaccade trials, the representation of the singleton decreases, and the representation of the location opposite the singleton, the endpoint of the antisaccade increases (indicated by the weaker spotlight on the singleton and growing spotlight on the saccade endpoint). At this same time in prosaccade trials, the representation of the saccade endpoint is enhanced by the selection that occurs in the Type II neurons (indicated by the highlighted spotlight on the singleton). Subsequently, in antisaccade trials, the endpoint of the saccade becomes selected more than the location of the singleton by Type I (thin, red, dashed) and Type II (thicker red, dashed) neurons (indicated by the highlighted spotlight on the antisaccade endpoint). The time taken to select the endpoint of the saccade predicts some of the delay and variability of RT. Modified from Sato and Schall [163].
References
-
- Kiorpes L. Visual processing in amblyopia: animal studies. Strabismus. 2006;14(1):3–10. - PubMed
-
- Economides JR, Adams DL, Jocson CM, Horton JC. Ocular motor behavior in macaques with surgical exotropia. Journal of Neurophysiology. 2007;98(6):3411–3422. - PubMed
-
- Lennerstrand G. Strabismus and eye muscle function. Acta Ophthalmologica Scandinavica. 2007;85(7):711–723. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
